筋トレの分子生物学

この稿では筋力トレーニングにおける筋細胞内の分子の挙動を述べます.参考にした書籍は以下ですが,最新の知見については専門家に譲ります.
Essential 細胞生物学は関連サイトが充実しており,学生向け資料のアニメーションやビデオ映像を見るだけでも価値があります.
1. 筋肉の発生
2. 筋細胞の構造
3. 物質輸送に関わる膜タンパク
4. 神経筋接合部における脱分極
5. アクチンとミオシン
6. ATPは細胞内の共通のエネルギー通貨である
7. 解糖系~細胞質でのATP産生機構
8. 電子伝達系~ミトコンドリアでのATP産生機構
9. クレアチンリン酸とクレアチンホスホキナーゼ
10. 筋肥大のメカニズム
11. メカニカルストレスとしてのエキセントリックコントラクション
12. 酸素負債では筋細胞内のATP産生のためミトコンドリアがフル稼働している
1. 筋肉の発生
受精卵が分裂を始めてしばらくすると,胚と呼ばれる細胞の塊になります.後に分化する器官ごとに内胚葉,中胚葉,外胚葉に分かれてきます.中胚葉から分化した脊索の伸長に伴い,脊索の両側に配置している中胚葉は体節を形成します.この中胚葉は板状の形をしており,椎骨,肋骨,分節化した筋肉を形成する基になります.つまり,筋肉は中胚葉から分化します.別の体節から体節細胞が側方の分節化していない中胚葉に移動し,四肢を含め体の骨格筋細胞のもとになります.この時期の細胞は筋芽細胞と呼ばれ,まだ分化が十分ではありません.筋芽細胞は衛星細胞として成人にも残っており,筋線維の修復の際に分裂して筋細胞を提供します.

©Wikipedia
筋芽細胞では筋特異的タンパク質はまだ十分に発現していません.最初のシグナルが加わるとまず筋芽細胞で MyoD ファミリー及び MEF2 ファミリーの 2 系統の筋形成遺伝子調節タンパクが発現します.次いで筋形成遺伝子調節タンパクは自身の遺伝子の転写を促進し,筋特異的タンパク質の遺伝子の転写を活性化します.このプロセスにはポジティブフィードバックがかかっており,最初のシグナルが消失しても持続します.
筋芽細胞は増殖を終えるとアクチン,ミオシン,トロポミオシン,トロポニン,クレアチンホスホキナーゼ,アセチルコリン受容体などの筋特異的タンパク質を発現します.これが筋芽細胞の分化です.筋芽細胞は分化するにつれて融合し,多核の筋細胞となります.筋芽細胞と筋線維は線維芽細胞の作る結合組織の網目構造の枠組みの中に保たれます.この枠組みが筋の発生を導き,筋細胞の並び方と向きを制御します.
筋肉の成長には筋線維の数,大きさ,長さ,太さが関与します.筋線維の長さは既存の筋線維への新たな筋芽細胞が補充されることで長くなります.筋細胞の数と大きさはミオスタチンにより抑制されます.ミオスタチンは骨格筋細胞から分泌される細胞外シグナルタンパクで,筋肉の成長制御に負のフィードバックを与えます.
筋細胞では活動依存シナプス除去が起こります.まず神経軸索末端と筋細胞の間でシナプスが形成されます.次いで筋細胞膜の接触場所ではアセチルコリン受容体が集まり,さらに軸索末端では神経伝達物質の分泌のための構造が組織されます.筋細胞は最初,複数の神経とシナプスを形成しますが,数週間経つと一つの神経接続のみが残ります.
2. 筋細胞の構造
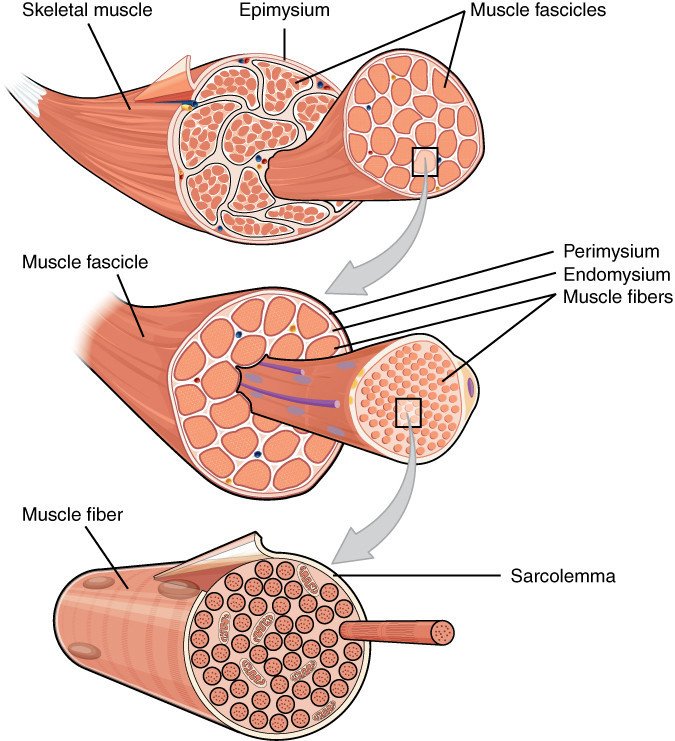
骨格筋細胞は随意運動を担い,長さは数 cm, 太さは 0.05 mm ほどで筋線維とも呼ばれます.骨格筋線維の数はヒトでは出生前に決まりますが,個々の線維細胞内の核の数は増減します.アスリートの筋肉の太さの増大には筋芽細胞の補充と筋原線維の大きさや数の増大が関わっています.
©Wikipedia
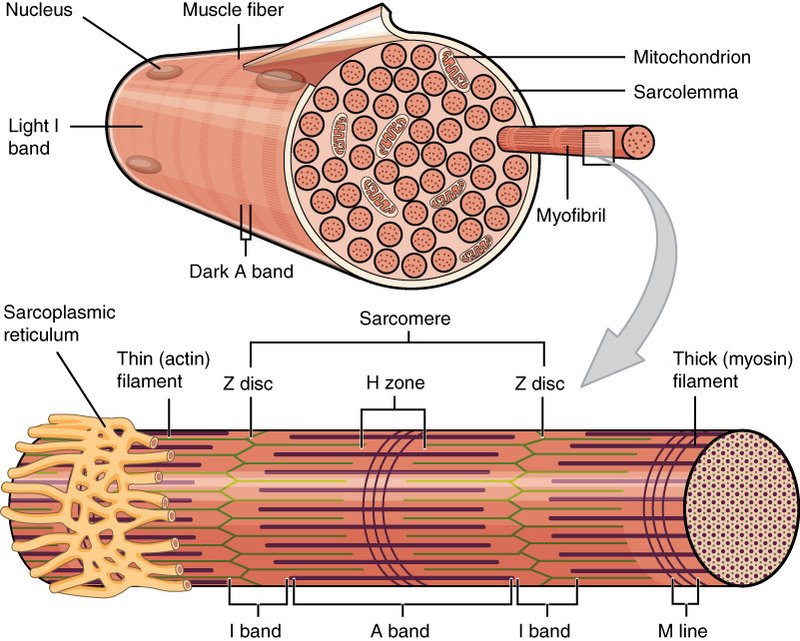
筋線維の細胞質の大半は筋原線維で占められています.筋原線維の直径は 1-2 µm で長さは筋線維とほぼ同じです.筋原線維は筋節(サルコメア)と呼ばれる収縮単位の反復構造からなり,その長さは 2.2 µm です.筋原線維を電子顕微鏡で観察すると,規則的な縞模様が見えます.明帯と暗帯はそれぞれアクチン,ミオシンに相当します.明帯の中央には Z 盤があり,Z 帯と Z 帯との間がサルコメアです.
©Wikipedia
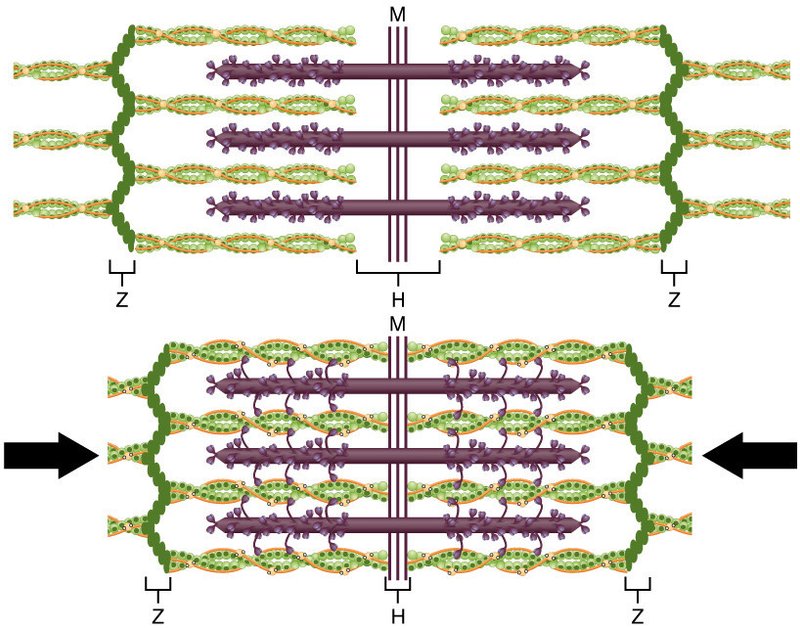
サルコメアは細いアクチンフィラメントと太いミオシンフィラメントが重なって規則的に配列している構造です.アクチンフィラメントは Z 帯に付着しており,ミオシンフィラメントの両端はタイチンというタンパク質を介して Z 帯に結合しており,タイチンはばねのように伸縮します.タイチンによってミオシンフィラメントはサルコメアの中央に固定されています.
©Wikipedia
ミオシンフィラメントは II 型ミオシン分子が 100 個以上集合してできています.II 型ミオシンは頭部を両端に向けて自己集合する性質があり,ミオシンフィラメントの集合した尾部の太い構造からミオシン頭部がフィラメントの外周に顔を出しています.
II 型ミオシンは 2 個の頭部と 1 本の長い尾部からなる双頭の蛇のような構造をしています.II 型ミオシン頭部にはそれぞれ ATP 結合部位があり,ミオシン頭部の先端にはアクチン結合部位があります.ミオシン頭部には ATP の加水分解エネルギーを頭部の機械的運動に変換するアクチュエーターとしての働きがあります.
サルコメアの断面を電子顕微鏡で観察すると,ミオシンフィラメントが六角格子状に配列し,その間にアクチンフィラメントが等間隔で配列しています.ミオシンフィラメントの断面は大きな点として見え,アクチンフィラメントの断面は小さな点として見えます.その数の比は 1:2 となっています.
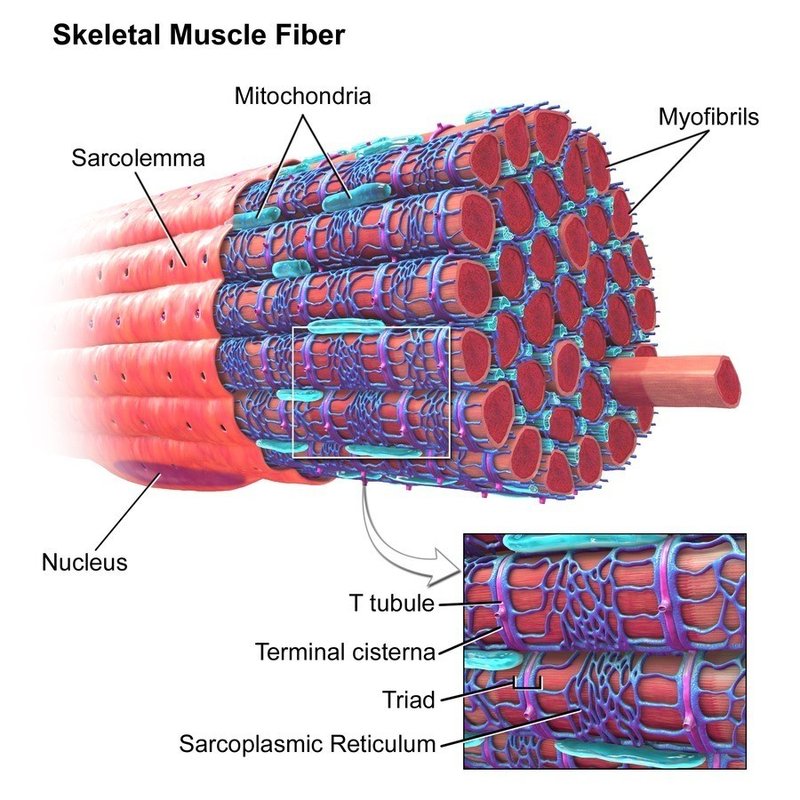
筋線維の細胞質には筋原線維の他に筋小胞体,ミトコンドリアがあります.また筋線維に特徴的な構造として T 管があります.筋小胞体は網目状に変形した小胞体で,筋原線維を網タイツのように包み込んでいます.筋小胞体の内部にはカルシウムイオン (Ca2+) が蓄えられています.ミトコンドリアはエネルギー産生の中心となる細胞内小器官です.電子顕微鏡で観察すると,ミトコンドリアは筋原線維に隣接して配置しています.T 管は筋細胞膜が内側に貫入してできたもので筋原線維を取り囲み,筋小胞体に隣接しています.
©Wikipedia
3. 物質輸送に関わる膜タンパク
ミトコンドリアの役割はアデノシン三リン酸 (ATP) を産生することで,産生された ATP は速やかに細胞質に運び出されます.細胞内の ATP 濃度は ADP の 10 倍以上に保たれています.
細胞膜は脂質二重層でできており,分子の大きさや疎水性の程度によって拡散・透過させやすさが異なります.一般には分子が小さく,脂溶性であるほど拡散速度は大きいとされています.酸素や二酸化炭素,窒素などの疎水性分子は非常に速く拡散します.水や尿素,グリセロールなどの小型で電荷を持たない極性の分子は速度は遅いものの脂質二重層を超えて拡散します.しかしプロトンやナトリウム,カリウム,カルシウム,重炭酸,塩化物イオンなどの電荷を持つ分子はどんなに小さくても脂質二重層を通り抜けることは困難です.グルコースやショ糖など,電荷を持たなくても大型の極性分子の透過速度も遅いです.これらのイオンや大型の分子を細胞膜を超えて運搬するにはエネルギーが必要です.
細胞膜や細胞内小器官の膜には物質輸送のためのタンパク質複合体が存在します.膜タンパクにはエネルギーを消費して物質を能動輸送するもの,何らかの刺激に応じてゲートが開き特定の物質を通過させるものがあります.前者は膜を隔てた濃度勾配に逆らって物質を輸送し,後者は膜を隔てた濃度勾配に従って物質を通過させます.前者を運搬体タンパク,後者をチャネルタンパクと呼びます.チャネルタンパクのゲートを開く刺激には膜電位の変化,伝達物質の結合,機械的刺激などがあります.チャネルタンパクのゲートには特定の物質のみを通過させるフィルター機構が備わっています.
細胞膜にある Na+/K+ ATP アーゼは運搬体タンパクで 1 個の ATP を ADP に加水分解するごとに 3 個のナトリウムイオン (Na+) を細胞外に汲み出し,2 個のカリウムイオン (K+) を細胞内に取り込んでいます.細胞内の Na+ 濃度は低くなり,細胞質体の有機分子のもつ負電荷を相殺するために別の陽イオンが必要になります.K+ がその役割を担います.その結果 K+ の細胞内濃度は細胞外よりも高くなり,Na+ はその逆になります.細胞内の負電荷に起因する陽イオンを細胞内に引き込む電気的吸引力と,濃度勾配に従って細胞外に流出しようとする力が釣り合って K+ は平衡状態にあり,膜電位が生じます.Na+ の濃度勾配は極めて重要で,栄養分を細胞内に取り込む殆どの輸送は Na+ の濃度勾配が駆動力となって行われます.また細胞内の pH 調節や浸透圧の調節にもこの Na+/K+ ATP アーゼが重要な役割を果たしています.細胞の消費するエネルギーの 1/3 はこのポンプを動かすのに消費され,神経細胞では 2/3 にもなります.
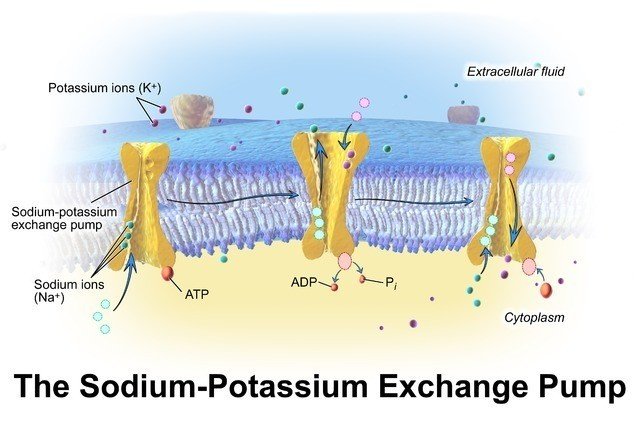
©Wikipedia
細胞外の Ca2+ 濃度は 10 mM ですが,細胞内の Ca2+ 濃度は 1 µM と低く保たれています.Ca2+ を細胞外に能動輸送する Ca2+ ポンプの働きによります.筋小胞体膜にも Ca2+ ポンプがあり,Ca2+ ATP アーゼと言い,これも運搬体タンパクの一つです.Ca2+ ATP アーゼは筋小胞体の膜タンパクの 90 % を占めており,1 分子の ATP を加水分解して 2 個の Ca2+ を細胞質から筋小胞体の内腔に能動輸送し,休止中の筋細胞の細胞質カルシウム濃度を低く保ちます.筋小胞体は Ca2+ の貯蔵所です.
神経細胞,筋細胞,内分泌細胞や卵細胞など電気的に興奮する細胞の細胞膜には電位依存性陽イオンチャネルがあります.特に神経細胞や骨格筋細胞には電位依存性 Na+ チャネルがあり,膜電位の負電荷が減少する脱分極の発生に関わっています.脱分極がある程度以上集積すると細胞膜全体に活動電位が発生します.電位依存性の Na+, K+, Ca2+ チャネルは膜貫通領域の一つに正電荷をもつアミノ酸を含んでおり,この領域は脱分極が起こると外側に移動してこれがチャネルの開口につながるコンフォメーション変化を起こします.
刺激により周辺で脱分極が発生すると電位依存性 Na+ チャネルが開き,Na+ が電気化学的勾配に従って流入します.これにより膜の脱分極が進み,さらに多くのチャネルが開いて膜の脱分極を進行させます.電位依存性 Na+ チャネルには自動的に働く不活性化機構があり,膜の脱分極が進行すると速やかにゲートが閉じます.ゲートが開いてから閉じるまでの時間は数分の 1 ミリ秒です.不活性化状態は数ミリ秒続きます.神経細胞には活性化した膜の電位を素早く元に戻すための電位依存性 K+ チャネルもあります.このチャネルは膜電位の変化にやや遅れてゲートが開き,Na+ 流入に続いて K+ の流出が起こります.その結果 Na+ チャネルが不活性化する前に膜電位は平衡電位に戻ります.
個々の電位依存性 Na+ チャネルは開くか閉じるかのいずれかの状態を取ります.全か無の方式といいます.チャネルがいつ開くかは決まっていませんが,開いた際には 1 ミリ秒あたり 1000 個以上のイオンが通過します.細胞膜を横切る電流の合計は個々のチャネルの開き具合ではなくその時点で開いているチャネルの総数を示します.これは筆者の推測ですが,脱分極開始を起点とした時間を横軸に,ゲートの開いたチャネル数を縦軸にプロットすると正規分布するのではないかと考えます.
活動電位を発生するのは Na+ チャネルだけではありません.筋細胞,卵細胞,内分泌腺細胞では電位依存性 Ca2+ チャネルが活動電位を発生させます.電位依存性陽イオンチャネルの構造や機能は多様ですが,それぞれのアミノ酸配列はよく似ており,進化的に近縁にあって設計原理が共通なのだろうと考えられています.
伝達物質依存性イオンチャネルの代表は骨格筋細胞のアセチルコリン受容体です.アセチルコリン受容体は神経筋接合部の筋細胞の細胞膜に高密度に分布しており,神経末端から放出されるアセチルコリンによって一過性に開き,陽イオンが通過します.電位依存性イオンチャネルと違って陽イオンの選択性は低く,Na+, K+ の他僅かに Ca2+ が通過しますが,実質的には Na+ が細胞内に流入します.この流入がシグナルとなって筋細胞膜の脱分極が始まり,筋収縮が起こります.
アセチルコリン受容体に結合したアセチルコリンはアセチルコリンエステラーゼにより加水分解されます.結合していた神経伝達物質がなくなると受容体は静止状態に戻りますが,アセチルコリンが受容体に結合したままだとチャネルが不活性化して脱感作状態となります.有機リン系の農薬はコリンエステラーゼ阻害作用があり,アセチルコリンが受容体に結合したままの状態を作り出して神経筋接合部の信号伝達を阻害して毒性を発揮します.
Action Potentials©Essential Cell Biology
ここから先は
¥ 252
この記事が気に入ったらサポートをしてみませんか?