激変する地球環境の下で生命は誕生し、進化してきた(遺伝子決定論は正しいか? ⑤)

地球環境の激変
ではなぜ、 生物はこれほどまでに手間暇かけて、遺伝的多様性を求めて来たのでしょうか。それは、激変する環境に適応して生き残る為であると考えられます。生息環境に変化が無いのなら、それに適応した遺伝子セットを一つ持っていれば十分です。しかし、何時どんな環境変化に見舞われるか予測不能の状況であれば、無駄になる可能性が有るにしても、遺伝的多様性を保持しておく事は生き残りの為には不可欠でしょう。実際、地球環境は、我々の日常感覚からは想像もできない程の、激変を繰り返してきたのです。地球規模での環境の激変は、生物の大量絶滅を繰り返し引き起こし、その度ごとに新たな生物が進化してきて、 崩壊した生態系を再建してきた訳です。生物進化の歴史は、繰り返す大量絶滅と生態系の再建の歴史だったのです。
そもそも地質年代の時代区分自体が、発見された動物化石の変化を基準に分けられています。下の図29)「顕生代の海洋生物の多様性」を見ても、地質年代の境目で大量絶滅が起こっている様子が分かると思います。
図29)顕生代の海洋生物の多様性(属レベル)
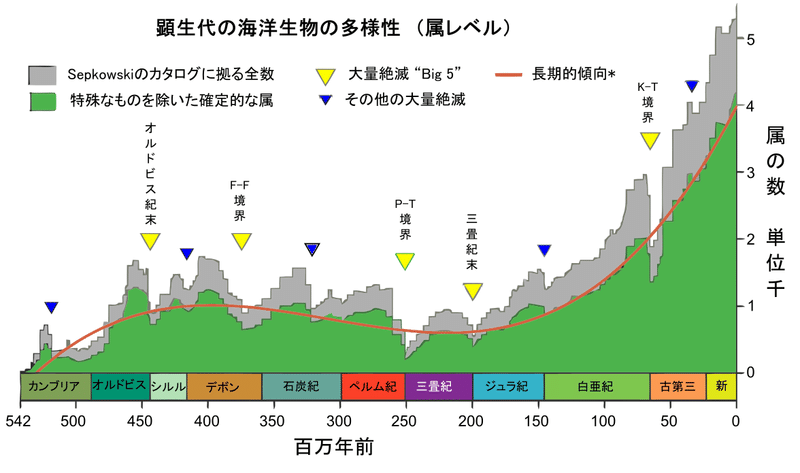
(出典:ウィキメディア・コモンズ )
煮えたぎる深海底で生命誕生
そもそも生命自身が、現在の穏やかな地球環境からは想像も出来ない程の、過酷な環境下で誕生しているのです。この事実は、生命とは何か、その本質を考える上で示唆に富んでいます。
地球が誕生したのは約46億年前、軌道上の無数の微惑星が衝突・集積して形成されました。地球誕生直後の45.5億年前には、火星ほどの大きさの天体が原始地球に衝突して、月が誕生しています(ジャイアント・インパクト説)。誕生直後の月は、地球との距離が1.5~2万キロ(現在は38万キロ)と異常に接近し、地球の自転速度も格段に早く、一日がわずか 5~8時間に過ぎなかったと言います。そして月が徐々に遠ざかるにつれて、地球の自転速度もそれに合わせて低下して行ったのです。
原始地球の直径が、現在の半分ほどに達した頃には、1年間に平均1000個以上の微惑星が落下していました。微惑星は毎秒数kmから十数kmという高速で原始地球に衝突し、その瞬間そこに含まれていた水や二酸化炭素は一瞬で蒸発し、放出されたガスは原始地球が月程度の大きさに達した頃から地表を覆い始め、次第に濃さを増しながら原始大気を形成して行ったのです。この原始大気の 80%は水蒸気で、残りは一酸化炭素から成っていたと考えられています。また、衝突した微惑星の運動エネルギーによって発生した大量の熱で、地球表面はドロドロに溶け、マグマの海(マグマオーシャン)に覆われていました。また、アポロ計画で持ち帰った月の石の分析結果から、約40億~38億年前には、集中的に大量の巨大隕石が月に落下した事が判明しています。そして同時期に地球にも、大量の隕石が落下したと考えられています(隕石重爆撃期)。
微惑星の衝突が徐々に収まり、表面温度が低下してくると、微惑星から大気中に放出されていた大量の水蒸気が、300度近い熱湯の雨となって降り注ぎ、原始の海が誕生します。最古の海の痕跡は、グリーンランドで発見された40億年前の花崗岩で、海洋プレートの沈み込み場所に生成したものと言われます。花崗岩質のマグマは、含水鉱物を含んだ海洋プレートが海溝からマントル内に沈み込み、融解する事で出来上がります。実は、花崗岩は地球独特の岩石で、海が形成されなかった他の惑星では、地表は基本的に玄武岩質岩石からなり、花崗岩は存在しないのです。
地球誕生から40億年前までは岩石自体が非常に稀で、地質学的証拠がほとんど無いこの時代を冥王代、 以後を太古代(始生代、40億~25億年前)と区分しています。38億年前には、海底に噴出した溶岩が固まって出来た枕状溶岩や、堆積岩も世界中で発見されています。最古の岩石は約40億年前のものですが、まとまった地層が世界各地で見つかるのも 38億年前からです。それ以前の地層が残っていないのは、当時の高温で活発なマントル対流によって、形成された地殻がすべてマントル内部に取り込まれてしまった為と考えられます。また38億年前には、プレートテクトニクスが機能していた事も分かっており、大陸地殻が海溝に沿って形成され始めていたと思われます。
現在の地表付近に残る炭素量から計算すると、海が誕生する以前の原始大気には、約60気圧もの二酸化炭素が含まれていたと言います。しかし海が誕生すると、大気中の大量の二酸化炭素は海水に溶け、カルシウムやマグネシウムの陽イオンと結合して炭酸塩となって沈殿して、気圧は数気圧にまで低下します。また温室効果の減少によって、気温も低下して行きました。しかし、大気圧が1気圧に下がるまでは、依然として海水温は100℃を超えたままでした。温泉や熱水噴出孔で生息する超好熱菌の中には、122度の高温でも増殖できる古細菌がいると言われます。この高温の海水温が、タンパク質やRNAが変性せずに安定して存在可能な100℃近くまで低下してくる中で、最初の生命が誕生するのです。
最古の生命の痕跡は、2017年に東京大学の研究チームが、カナダ北東部の北ラブラドル地域のサグレック岩体で発見した、約39億5000万年前の最古の変成堆積岩に含まれる炭質物(グラファイト)です。この炭質物は、数十〜数百μmの棒状や球状で、炭素同位体の組成から、代謝経路を経て生じた生物起源のものであるとされています。このことは、生命誕生が約40億年前の海の誕生とほぼ同時期に起こっていた事を示しています。つまり生命は、長い試行錯誤の後に奇跡的に誕生したものなどでは無く、生命活動が可能な地球環境が生まれるや否や、ごく短期間で出現していたのです。これは、生命誕生が奇跡的な偶然の産物などではなく、地球進化と分かち難く結びついた、必然的過程であった事を強く示唆しています。
しかも、海が出現した頃の地球は、地下のドロドロに溶けたマグマの層と、その表面を覆う薄い原始地殻の上に150℃近くの熱湯の海が一面に広がり、それらを二酸化炭素の大気が包むという成層構造を形成していたと考えられます。そしてこの熱湯の海には、まだ大量の隕石が降り注ぎ、すぐそばを公転する月の強力な潮汐力が、海水を烈しくかき混ぜていました。大量の熱を蓄え、あちこちで海底火山が噴火し、激しく活動していた形成途中の原始地球の煮えたぎる熱湯の海の中で、最初の生命は誕生したのです。
熱水噴出孔と生命
実は、生命が誕生した原始の熱湯の海に似た環境が現在も存在しています。それは深海底の熱水噴出孔です。1977年、アメリカの深海調査艇アルビン号は、ガラパゴス諸島の北東320 km、深度2600mの深海底で、高温の熱水が噴出する熱水噴出孔(ハイドロサーマルベンド)を発見しました。しかも驚くべき事に、この太陽の光さえ届かない高温・高圧の極限環境に、高密度の特異な生物群落が存在し、砂漠のオアシスの様に、暗黒の深海底で熱水噴出孔の周りに特異な生態系を形成していたのです。熱水噴出孔は、中央海嶺の様な海洋プレートの湧き出し口、海底火山、火山列島の様な島孤の後ろ側にできる窪んだ海域(背孤海盆)などで発見されています。日本付近でも、那覇市の北北西130 kmの伊是名海穴、さらに北へ40 km行った伊平屋海嶺などが知られており、ここからは200℃を越える熱水が噴出しています。
図28)熱水噴出孔(ブラックスモーカー)
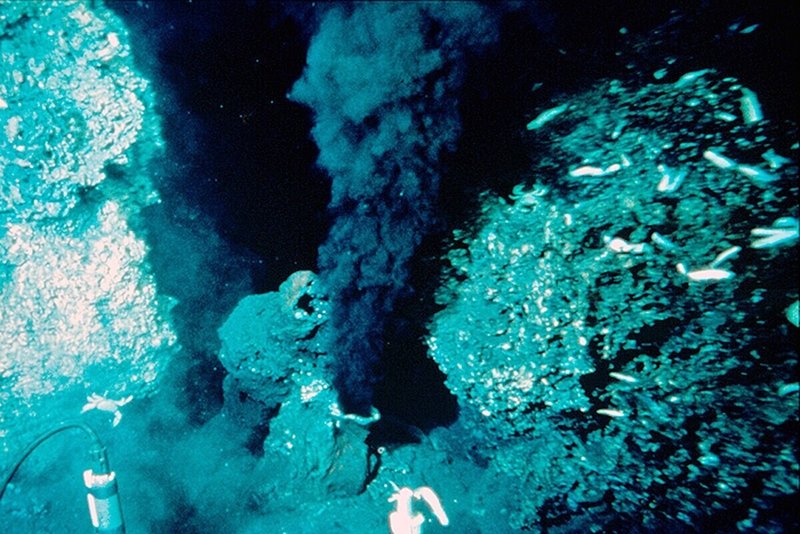
(出典:ウィキメディア・コモンズ )
熱水噴出孔は、地殻の直下まで迫ったマグマ溜りの周辺に染み込んだ海水がマグマによって熱せられ、大量の金属イオンを含む高温・高圧の熱水となって海底から噴出したものです。熱水は上昇して来る時に周囲の玄武岩と反応して強い酸性となり、そしてケイ酸塩鉱物からは大量の金属イオンを溶かし出して来ます。さらに、熱水中に含まれる硫酸イオンが玄武岩中の二価の鉄イオンによって還元され、大量の硫化水素が発生しています。熱水中の金属イオンはこの硫化水素と反応し、硫化物の形で上昇する通路の壁や噴出口付近に沈殿して行きます。こうして、噴出口の周りには、析出した金属によって煙突構造(チムニー)が形成され、その先端から大量の金属イオンや硫化水素を含む熱水が、黒煙の様に立ち上る様になるのです。これがブラック・スモーカーです。アルビン号によって発見された熱水噴出孔の温度は 350℃に達し、その周囲の半径数百mに渡って水温2~50℃の局所的な温水域、ホット・スポットができていました。そこに、高密度の熱水生態系が形成されていたのです。ここでは、熱水中の硫化水素を酸化してエネルギーを取り出して炭素同化を行う、化学合成細菌のイオウ酸化バクテリアが一次生産者となり、このバクテリアを体内に大量に共生させた大型の二枚貝や管状のチューブ・ワーム、それらを餌にするエビやカニなどの甲殻類が多数生息しています。この深海で最も繁栄しているチューブ・ワームは、口や肛門・消化管もなく、ただ硫化水素を鰓で取り込んで共生バクテリアに供給し、それが作り出すエネルギーや栄養素に依存して生活しているのです。その体内に共生するイオウ酸化バクテリアの量は体重の90%にもなると言います。
図29)チューブ・ワーム

(出典:ウィキメディア・コモンズ )
オレゴン州立大学のJ.コーリスらは、このホット・スポットが太陽エネルギーとは無縁の環境下で、特異な生態系を成立させている点に注目して、深海底の熱水噴出孔で生命は誕生したという仮説を発表しました。以来、この熱水噴出孔説は、生命誕生の最も有力な候補の1つになっています。熱水噴出孔の特徴は、第1に高温・高圧な事で、ここには化学反応を進めるのに必要な熱エネルギーがふんだんに存在します。岩の裂け目からしみ込んだ海水は、マグマの熱で急激に熱せられ様々な化学物質を取り込みますが、マグマとの接触面を離れると今度は周囲の海水によって急速に冷却されるため、高温で生成した物質は熱による分解を避ける事ができます。これは実験室で様々な分子を作る方法と同じで、熱水噴出孔はいわば熱勾配のついた一種の流動性の反応炉となっているのです。第2の特徴は、メタン・水素・硫化水素・アンモニアなどの還元性ガスの濃度が極めて高く、周りの海水よりも還元的な環境になっている事です。一般的に、酸化的なガスからは有機物は出来難いが、還元的なガスからは容易に生成する事が分かっています。つまり熱水噴出孔は、有機物の生成し易い環境になっているのです。事実、紅海の熱水噴出孔の海水からは、アミノ酸のグリシンが周囲の海水よりも10倍も高い濃度で検出されると言います。これは、熱水噴出孔の周辺で、有機物が無生物的に合成されている可能性を示唆しています。さらに第3の特徴は、鉄・マンガン・銅・亜鉛などの金属イオンの濃度が、通常の海水の1000倍以上と極めて高い事です。これらの金属イオンが、熱水中での有機物合成反応に関わり、生命誕生に重要な役割を果たした可能性が考えられるのです。
硫化水素存在下でのペプチド合成
熱水噴出孔からは硫化水素が大量に放出されている訳ですが、実はこの硫化水素が生命誕生の鍵を握っていた可能性があります。しかも、深海の熱水噴出孔の様な高温・高圧の特殊な環境でなくても、硫化水素の存在下で有機分子、それもアミノ酸ばかりでなくペプチド(2個以上のアミノ酸がペプチド結合した短鎖分子。50個以上はタンパク質)や糖までが、極めて容易に自然環境の中で現在も生成されている様なのです。そして、この有機分子生成のカギが、熱水噴出孔でも大量に存在する硫化水素だったのです。
それは1986年、京都大学の藤永太一郎と海洋化学研究所の紀本岳志によって、日本最大の汽水湖である中ノ海の環境問題に関連した、水質成分の連続計測から偶然に発見されました。汽水湖というのは、海水と淡水が入り込む海に連なる湖で、比重差から湖底に海水が溜り、その上に河川水が乗って成層湖となったものです。中ノ海は、宍道湖から流れ込む淡水と日本海からの海水が一年中混じり合う事なく、表層の溶存酸素濃度の高い酸化性の淡水と、底層の貧酸素で還元性の海水が常時密度成層をなしています。この湖では、深度6.5mの極貧酸素状態の湖底から硫化水素とアンモニアが大量に発生しているのですが、各種栄養塩類の垂直分布の測定から、上昇するに従いアンモニアが急減する事実が明らかになったのです。そこで、底層水中の浮遊粒子を加水分解して分析すると、アンモニア態窒素の減少に見合うアミノ酸濃度の増加が見出されたのです。つまり、ここではアンモニアからペプチドが無機的に合成されていたのです。
実験室で炭酸(ホルムアルデヒドやギ酸を使っても同じ)とアンモニアの混合溶液に、常温・常圧下で硫化水素ガスを通気する実験を行うと、それまで考えられた事もない反応によって、各種のアミノ酸構成を持つペプチドや糖(炭水化物)が生成されたと言うのです。そして、硫化水素の通気開始後わずか数時間で、ペプチドや糖が直径数 μmの大きさの揃ったコロイド状粒子となって溶液の底に沈殿しました。最初に、最も単純なアミノ酸のグリシン・モノマーが生成され、時間の経過と共にそのグリシンが重合してペプチドを作り、次第に溶解度を減少させて乳白色の沈殿が溶液中に生成したのです。しかも、この沈殿したコロイド粒子を顕微鏡で観察すると、溶液との界面で、活発に物質交換をしている事が分かりました。重合度の低いペプチドが表面から離れる一方、重合度の大きいものが粒子の表面に重合析出して行くといった、界面での代謝的反応が進行していたのです。こうして粒子は成長し、ある大きさに成長した所で溶解と析出が釣り合い、粒度の揃ったコロイド粒子が形成されたのです。
図30)硫化水素存在下での糖・ペプチドの自然合成
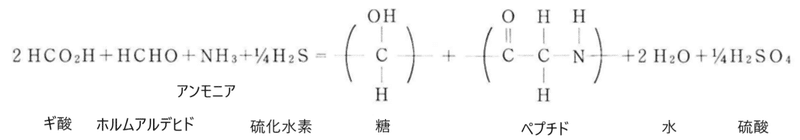
(奈良教育大学紀要第37巻第2号昭和63年「生命起源に関する新しい知見について」より改変)
ここで起こっている反応は、基本的には炭酸が還元されて各種のアルデヒドが生成し、これにアンモニアが加わってさらに還元され、多様なペプチド類が合成されるというものです。そして、この反応になくてはならない還元剤として硫化水素が関わっていたのです。
生命を形作る、炭水化物・脂質・タンパク質・核酸などの生体高分子はすべて炭素化合物で、炭素(C)に水素(H)・酸素(O)・窒素(N)が共有結合したものが、鎖状あるいは環状に連なり巨大分子を形成したものです。生命は、この巨大分子の骨組みの炭素を、二酸化炭素(CO2)やそれが水に溶けた炭酸(H2CO3)から得ています。つまり生物は、二酸化炭素に水素を付加し、酸素を引き抜いて還元する事で、有機分子を合成しているのです。
そして生体高分子は、それを構成する分子から水1分子が取れて、脱水縮合する事で合成されます。例えば、炭水化物の多糖類は、単糖類が脱水縮合して、鎖状にあるいは枝分かれしながら長くつながったものです。2分子の単糖が結合する時に1分子の水(H2O)が放出されて、単糖間に共有結合ができるのです(グリコシド結合)。タンパク質も同様に、20種類のアミノ酸が脱水縮合して(ペプチド結合)、1本の鎖状に長く連なったものです。脂肪や油、細胞膜を構成するリン脂質は、グリセロールと脂肪酸が脱水縮合して結合しています(エステル結合)。また遺伝子の本体である核酸(DNA・RNA)も、ヌクレオチドが脱水縮合して(ホスホジエステル結合)長く連なった多量体です。
このように生体高分子を作るには、還元反応と脱水反応が不可欠なのです。ところが硫化水素(H2S)は、カルボニル化合物などから還元反応で酸素を引き抜いた時に生じる無水硫酸が脱水反応にも働き、還元反応と脱水反応を同時に起こす事ができる、生体高分子の合成には打って付けの還元剤となっているのです。このような2重の働きをする還元剤は、他には知られていないと言います。つまり生命誕生のカギは、硫化水素にあったと言っても良いのです。
図31)硫化水素による還元作用と脱水作用
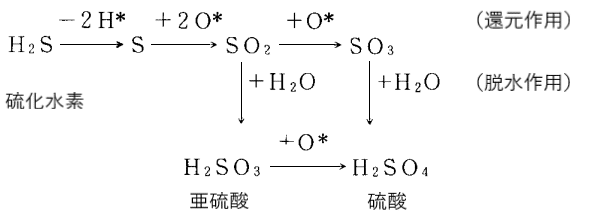
(奈良教育大学紀要第37巻第2号昭和63年「生命起源に関する新しい知見について」より改変)
先に、 RNA ワールドについて紹介しましたが、 有機分子が硫化水素の大量に存在する熱水噴出孔で容易に自然合成されるとなると、最初の生命はペプチドと RNA の相互作用の中で誕生したという、より現実的なシナリオが 浮かび上がってきます。
シアノバクテリアによる酸素放出
地球環境の変化には、生物自身も深く関わっています。その典型的な例が、シアノバクテリアによる酸素の放出です。この広範囲に分布する光合成細菌には、結合して糸状の多細胞の鎖を形成するものがあり、それが原始的な海藻(藻類)に似ている事から、かってはラン藻と呼ばれていました。しかし本当の藻類は真核生物であり、今日では混乱を避ける為にシアノバクテリアと呼ばれています。実は、酸素は大変反応性の高い元素で、直ぐに他の元素と化合して、二酸化炭素・水 ・その他酸化物となる為に、生命誕生以前には遊離した酸素分子は存在していません。現在の大気中に 21%含まれる酸素のほぼ全ては、光合成によって生み出されたものなのです。そして、地球で初めて光合成により、遊離酸素を作り出したのがシアノバクテリアだったのです。またシアノバクテリアは、真核細胞に細胞内共生して葉緑体を誕生させました。つまり、植物が行う酸素発生型の光合成は、シアノバクテリアに起源する同じものなのです。
図32) 糸状性のシアノバクテリア
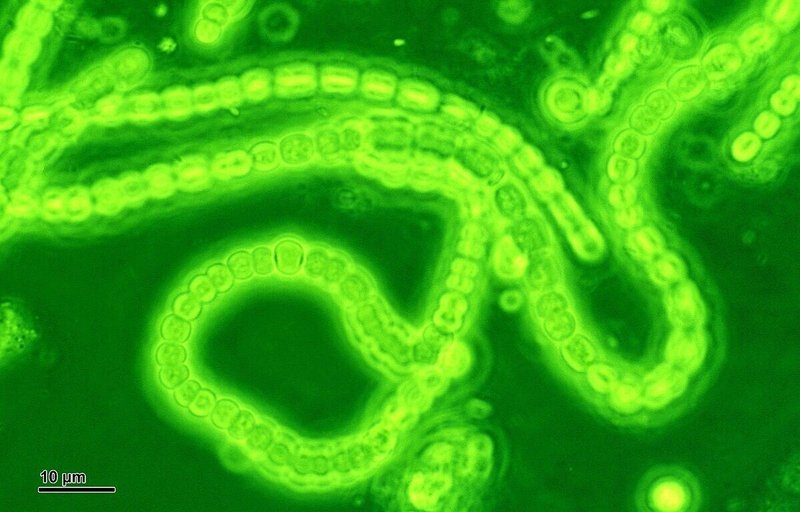
(出典:ウィキメディア・コモンズ )
光合成細菌の中でも、酸素を放出できるのはシアノバクテリアだけです。 これはシアノバクテリアが、光合成細菌の中でも唯一、光化学系Ⅰと光化学系Ⅱという2つの光化学系を持つ事に起因しています。葉緑体やシアノバクテリアでは、光化学系ⅡのP680反応中心(680nm付近の光を吸収する)のクロロフィルa分子に光が当たると、電子はその光量子のエネルギーを吸収して高いエネルギー状態に励起され軌道を変えます。こうして活性化された電子は、反応中心のクロロフィルaを離れて電子伝達系に渡され、伝達体から伝達体へと移動する間にプロトンポンプを駆動してプロトンをチラコイド内腔に汲み上げ、最終的に光化学系Ⅰに入ります。この過程で、電子を失い強い酸化力を獲得した光化学系ⅡのP680は、不足した電子を水から奪ってこれを酸化し、水をプロトンと酸素に分解して酸素分子を発生させるのです。光化学系ⅠのP700で再び光エネルギーを吸収して励起された電子は、ストロマに存在するNADP+に渡され、これを還元してNADPHを生成します。このNADPHが、二酸化炭素を還元して糖を合成するのです。また、水の酸化と電子伝達系でチラコイド内腔に汲み出されたプロトンは、 ATP 合成に利用され、炭素固定にエネルギーを供給する事になります。
図33)葉緑体のチラコイド膜での光化学反応と電子伝達系
(PSⅠ:光化学系Ⅰ、PSⅡ:光化学系Ⅱ、e-:電子)
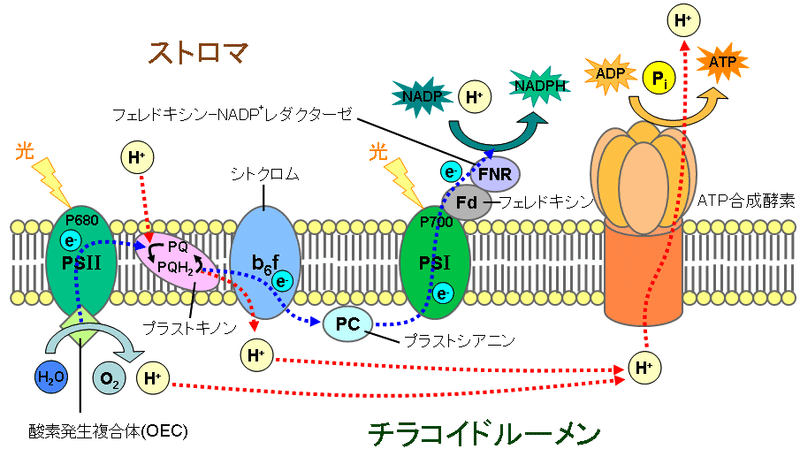
(出典:ウィキメディア・コモンズ )
図34)葉緑体とシアノバクテリア
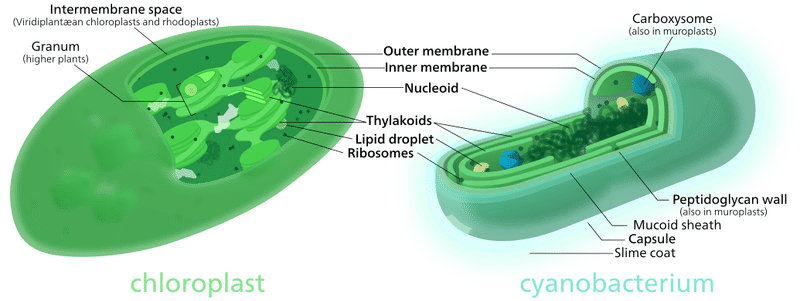
(出典:ウィキメディア・コモンズ )
シアノバクテリア以外の光合成細菌は、光化学系Ⅰと光化学系Ⅱのどちらか一方しか持たず、水の光分解は起こりません。水の代わりに、電子供与体として硫化水素などを利用して、酸素の代わりに硫黄などを析出しています。 シアノバクテリアは、光化学系Ⅰ・Ⅱの二つの光化学系を持つ唯一の光合成細菌な訳ですが、緑色硫黄細菌の光化学系Ⅰ、紅色細菌類の光化学系Ⅱの遺伝子が水平移動して、二つの光化学系を合わせ持つシアノバクテリアが誕生した可能性が考えられます。 つまり、異種の生物間で遺伝子を交換し、 融通し合う事で新たな生物が進化した可能性が高いのです。
図35)光合成の収支式
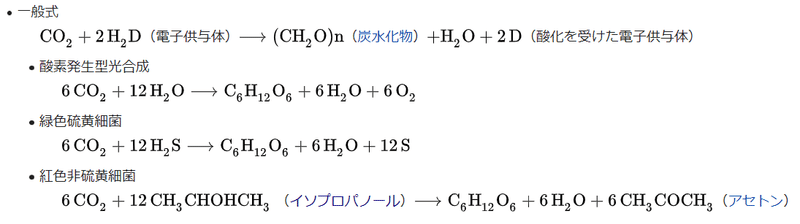
(出典:ウィキペディア )
2種類の光化学系を持つシアノバクテリアの登場は、地球生命の歴史に革命をもたらす事になります。光化学系ⅠとⅡを連結する事で、二酸化炭素を還元する為の水素源をそれまでの硫化水素から水に切り換える事が可能となり、その結果、光合成の廃棄物として大量の酸素が発生する事になったのです。この、シアノバクテリアにより大量に供給された酸素が、地球環境を激変させ、生態系を作り変えて行く事になります。
1種類の光化学系しか持たない光合成細菌では、水素源の硫化水素の発生場所が限られている為に、生態的分布を広げる事が困難でした。しかし、地球上に無尽蔵に存在する水を、水素源にする事に成功したシアノバクテリアは、水の惑星の中で爆発的に分布を広げる事に成功するのです。二酸化炭素・水・太陽光線さえあれば、何処でも生きて行けるシアノバクテリアは、太陽エネルギーを使って二酸化炭素・水といった地球の構成要素を有機物に変え、大量の有機物と酸素を供給して行く事になります。これによって、地球はそれまでの還元的環境から酸化的環境へと激変し、シアノバクテリアが作り出す大量の有機物を酸素呼吸によって二酸化炭素と水に分解して、効率的にエネルギーを生産する好気性従属栄養生物が進化してくる事になります。それが、我々の遠い祖先の真核細胞なのです。今日、私たちが直接目で見る事のできる生物のほとんどすべてが真核生物です。この豊かで多様性に富む生態系と真核生物の繁栄の基礎は、約27億~19億年前に起こった、シアノバクテリアの爆発的な大繁殖に淵源を求める事が出来るのです。
しかし、もっと根本的な変化は、 それまで地球内部からの化学エネルギーに支えられて誕生した地球生態系が、太陽エネルギーを基礎とする生態系に大転換した事です。それによって、無尽蔵とも言える莫大な太陽エネルギーを使って、膨大な量の有機物を合成する事が可能となり、その基礎の上に豊かで多様な生態系が誕生するのです。至る所に生命が満ち溢れる、今日の地球生物の繁栄の基礎は、シアノバクテリアが進化させた光合成によって築かれたのです。石油や石炭の由来が、植物や藻などの埋没した遺骸である事を考えれば、如何に膨大な量の有機物が合成されて来たか分かるでしょう。二酸化炭素と水を使った光合成に成功したシアノバクテリアの登場によって、地球は真の意味での生命の惑星となったのです。
シアノバクテリアは、37億年前に地球上に登場してから生き続け、しかもその構造は当時とほとんど変わっていないと言います。そして今日でも、海水・真水を問わず地球上の至る所で繁殖し、彼等の仲間は 150属1500種にも及びます。この意味でシアノバクテリアは、生命史上最も成功した生物と言う事もできます。そして、この生命進化の大傑作であるシアノバクテリアの光合成は、葉緑体としてそのまま高等植物にまで伝えられて来たのです。
図36)オーストラリア・シャーク湾のストロマトライト

(出典:ウィキメディア・コモンズ )
シアノバクテリアと言うと、オーストラリア・シャーク湾のストロマトライトを思い浮かべる人も多いでしょう。 これは、シアノバクテリアが形成する微生物マットの出す粘液が 、光合成が停止した夜間に、海水中に浮遊する微細なミネラルの粒子を捕らえて炭酸カルシウムと結合させ、毎日一層ずつ堆積させて上へ上へと成長して出来たドーム状の岩石です。
最古のストロマトライトとされるものは、西グリーンランドのイスア地域で、37 億年前のストロマトライト様の模様を示す堆積岩が発見されています。また、西オーストラリアのピルバラ地域でも、35 億年前のシアノバクテリアのマットが岩石化したものが見つかっています。しかし、生物起源を疑う意見もあり、シアノバクテリアの出現年代は良く分かっていません。ところが、シアノバクテリアの繁栄と、大量の酸素放出を示す間接的な証拠が存在しています。それが全世界に分布する縞状鉄鉱床です。
縞状鉄鉱床の形成
縞状鉄鉱床は、先カンブリア時代の海底に堆積した酸化鉄を主体とする堆積鉱床で、鉄分の少ないケイ酸塩鉱物のチャート(シリカの細粒形態)の層と、酸化鉄・磁鉄鉱・赤鉄鉱の薄い層が縞状に細かく互層をなし、非常に大規模な鉱床を形成したものです。例えば、北アメリカ大陸の鉱床では、カナダ東部のラブラドールからケベックにかけて1,000km、さらにアメリカ合衆国の五大湖周辺まで途切れながらも約2,000km以上も続いています。また、オーストラリア、ブラジル、カナダ、アメリカ、インド、ロシア、南アフリカ、ウクライナ、中国など、世界中に巨大な鉱床が存在し、現在採掘される鉄鉱石のほとんどがこの縞状鉄鉱床からのもので、世界の鉄埋蔵量の60%以上を占めると言われます。
図37)縞状鉄鉱床(西オーストラリア・ピルバラ)

(出典:ウィキメディア・コモンズ )
図38)西オーストラリア・ハマーズリー山脈の鉄鉱石鉱山

(出典:ウィキメディア・コモンズ )
また、戦前日本の国策会社の満鉄(南満州鉄道)が、遼東半島付け根の鞍山付近で発見し、鞍山製鉄所を設立して開発した大鉱脈も縞状鉄鉱床です。鞍山の鉄鉱石の埋蔵量は約100億トンで、中国全土の4分の1を占めるとされます。後に、昭和製鋼所と改称したその総生産能力は、1942年までには年間360万トンに達し、世界の主要な鉄鋼センターの1つになっていました。しかし戦後、ソ連は昭和製鋼所の設備を解体して自国に持ち帰っています。つまり、スターリンは戦利品として略奪して行った訳です。
図39)鞍山市の斉大山露天掘り鉄鉱石鉱山

(出典:ウィキメディア・コモンズ )
このように世界中に散在する巨大な縞状鉄鉱床ですが、実はその形成時期のほとんど全てが、先カンブリア時代の始生代と古原生代で、38億~19億年前の年代の地層に限られています。それ以後は、約7億年前のスノーボールアース直後の一時期に堆積しただけなのです。その中でも、27億年~19億年前に、非常に大規模な鉱床が集中的に形成されています。
図38)縞状鉄鉱床の形成(淡黄色:古い始生代の地層、濃黄色:ゴンドワナ大陸での形成、茶色:粒状の鉄鉱層、赤:スノーボールアース直後の形成、横軸:10億年前)
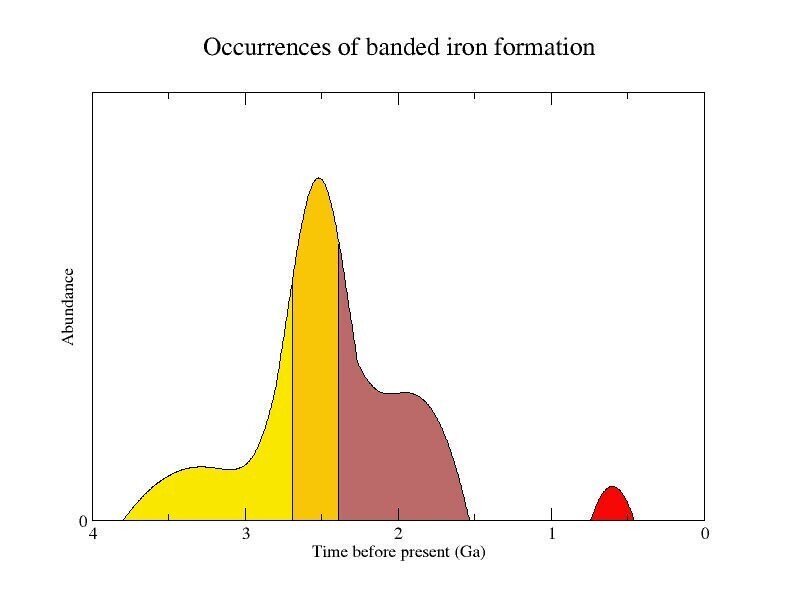
(出典:ウィキメディア・コモンズ )
この縞状鉄鉱床の集中的な堆積こそが、シアノバクテリアの大繁栄の証拠なのです。二酸化炭素を還元する水素を、水から得る事に成功したシアノバクテリアは、生息場所の制約から開放され、世界中にその分布を広げて行きました。そして、太陽光の良く届く浅海で大繁殖をします。この地球最初の人口爆発とも言うべき当時の大繁殖の跡は、ハマースレー渓谷一面に厚く積もったブラックシェールとして残っています。そして、シアノバクテリアの放出した大量の酸素が、世界各地に残る縞状鉄鉱床の巨大鉱床を形成する事になるのです。縞状鉄鉱床は、海水中に溶けていた2価の鉄イオンが、シアノバクテリアの放出する酸素によって酸化され、不溶性酸化鉄として沈殿して形成されたものなのです。
大酸化イベントとスノーボールアース(全球凍結)
シアノバクテリアの大繁殖は海中に大量の酸素を供給し、 世界各地に大規模な縞状鉄鉱床を誕生させた訳ですが、19億年前になると海中に溶けていた2価の鉄イオンもほとんどが沈殿して、縞状鉄鉱床の堆積も終わりを迎えます。約25億年前にピークに達した縞状鉄鉱床の形成は、19億年前以降の地質記録から大部分が消えてしまうのです。以後は、鉄イオンの酸化に費消されていた酸素は、海中から大気中に放出される事になります。その結果、以前にはほとんど存在していなかった大気中の酸素が、約24.5億~20億年前の原生代初期に、現在の1/100レベルにまで急上昇したと考えられているのです。そして、この爆発的な酸素濃度の上昇を、「大酸化イベント」と呼んでいます。しかも、酸素濃度は大酸化イベントで一旦、現在と同レベルにまで急上昇した後、約1億年かけて徐々に現在の1/100にまで低下した様なのです。
図33)大気中の酸素濃度の推移
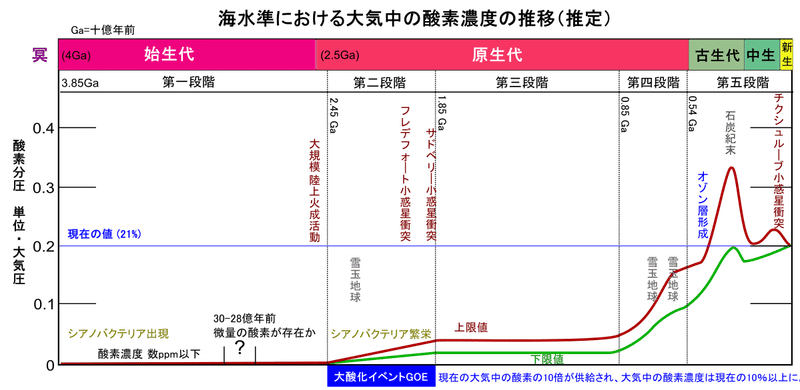
(出典:ウィキメディア・コモンズ )
実はこの時期、地球は深刻な氷河期に襲われています。原生代初期、約24.5億~22億年前のヒューロニアン氷河時代で、その最終期には地球表面全体が凍結するほど、厳しい寒冷化に見舞われていたのです。地球を覆う氷床は、太陽光を反射して寒冷化を加速し、最盛期には厚さ約3000mにも及ぶ氷床が全地球を覆っていたと言われます。これを、スノーボールアース仮説と呼んでいます。このヒューロニアン氷河時代を生む事になった原因は、シアノバクテリアにより大量に放出された酸素による、二酸化炭素やメタンなどの大気中の温室効果ガスの減少と考えられています。
図34)ヒューロニアン氷河時代のスノーボールアース

(出典:ウィキメディア・コモンズ )
また、全球凍結を終わらせる事になる原因も、火山活動により供給された二酸化炭素です。氷床の存在により、海水中に溶け込む事が出来ず大気中に蓄積した大量の二酸化炭素の温室効果により、氷床は数百年といった短期間で一気に解凍し、消滅して行ったと考えられています。そして、22億年前に全球凍結が融解した直後に、酸素濃度が現在のレベルにまで急上昇していたのです。
氷床が融解すると、今度は大量の二酸化炭素の強力な温室効果により、地球全体が一気に50℃近い高温環境になったとされます。こうなると大陸の風化作用の再開で、海洋にリンなどの栄養塩が大量に供給され、海洋の富栄養化が進行します。その結果、温暖な海洋表層においてシアノバクテリアが爆発的に大繁殖し、大量の酸素が一気に放出される事になったのです。それまで、現在の1/100000以下のレベルで安定していた酸素濃度は、現在とほぼ同レベルにまで急激に上昇し、その後、激増した酸素は、約1億年かけて徐々に減少して、現在の1/100のレベルで安定するのです。
「大酸化イベント」での、こうした急激な酸素濃度の上昇は、嫌気性細菌の大量絶滅を引き起こしたと考えられています。酸素は、生物にとっては極めて毒性の強い元素です。 生体分子は、酸化されると構造や性質が不可逆的に変化してしまうのです。 嫌気性細菌のほとんどは、酸素に触れるとすぐに死んでしまいます。元々、有機分子は二酸化炭素を還元する事で合成されます。 反対に、有機分子を酸化すると、最終的には二酸化炭素と水に分解されてしまいます。酸素の中で生きて行くには、その悪影響から細胞を守る防御機構を進化させる必要があったのです。
嫌気性細菌の多くが絶滅した一方で、それと入れ替わる様に新しい主役が登場して来ます。それが、我々の直系の祖先である真核細胞です。サイズが原核細胞の10倍(体積では1000倍以上)にもなる真核細胞が、好気的代謝に必要十分な酸素を細胞内に取り入れるには、酸素濃度が現在の1%(パスツール点)を超える事が必要と言われており、大酸化イベントでその条件が達成された訳です。 実際、最古の真核生物として、北米のスペリオル湖(五大湖の1つ)南岸の21億年前の縞状鉄鉱層の中から、真核細胞らしい化石が発見されています。幅1mm、長さ9cmのリボンがまるまった様な形のグリパニアで、この化石は中国やインドの11億年前の地層から既に発見されており、現生の真核藻類に良く似ていると言います。
図35)グリパニア

(出典:ウィキメディア・コモンズ )
全球凍結直後の多細胞生物の進化
ヒューロニアン氷河時代の全球凍結が終わった直後の約22億年前、シアノバクテリアの大繁殖によって酸素濃度が急上昇し、それに合わせるようにして真核細胞が登場して来ました。実はそれと同様の事が、新原生代クライオジェニアンの約7.2億年~6.35億年前、スターチアン氷河時代・マリノアン氷河時代の全球凍結の直後にも起こっていた様なのです。約6億年前、氷床が溶け地球が急速に温暖化する中で、酸素濃度が再び現在のレベルにまで急上昇し、その中で様々な多細胞生物が急速に進化して来たのです。
図36)エディアカラ生物群

(出典:ウィキメディア・コモンズ )
それが、南オーストラリアのアデレード北方、エディアカラ丘陵の浅海~潮間帯の堆積物から最初に発見された、不思議な生物化石のエディアカラ生物群です。これは、新原生代末の約6億~5.4億年前に繁栄した最古の多細胞生物で、オーストラリア中西部・南西アフリカ・イングランド・スウェーデン北西部・ウクライナ・モスクワ北東域・北シベリア・イラン・インド・中国・ニューファンドランド西部・アメリカ東部など世界各地から発見されています。その特徴は、硬い殻や骨格が無く、柔組織だけのクラゲの様に薄く柔らかい体の生物で、木の葉の様な形のものや三つ巴の模様のものなど、キルティングの様な薄くペラペラなシート状の軟体性の生物だった事です。しかも、サイズがかなり大きく、中には体長が1mを越す巨大なものまでいました。次の時代の、カンブリア紀のバージェス動物群や澄江動物群の多くが数cm程度であるのに対して、クラゲ状のものや楕円形の「ディッキンソニア」など、直径数十cmにもなる多種多様なシート状の生物が繁栄していたのです。
図37)ディッキンソニア

(出典:ウィキメディア・コモンズ )
その大きさに反して、エディアカラ生物には体を支える骨格や口・肛門をはじめ体内器官の痕跡が全く認められず、その多くは運動能力のない底生生物で、現生生物中に対応する仲間も見当たらない、動物なのか植物か、あるいはどのように生活していたのか全く謎の生物群なのです。実際、エディアカラの化石には死ぬ前に動いた痕跡がまったく認められず、動物にしては動きがなさすぎると言います。エディアカラ生物を、現生の生物群やその祖先に当てはめようという試みは、環形動物や刺胞動物など一部の少数例を除いて成功していません。彼等は、肺や腸そして心臓などの循環器官系を進化させる事なく、表面積を増して体のサイズを大きくする事に成功した生物で、従来の生物とは異なった系統の生物群とも考えられるのです。そして、シート状の表面積の大きな体制は、体内に大量の光合成藻類や化学合成細菌などを共生させる為のもので、エディアカラ生物はそれらの微生物に生活の場を提供する宿主であった可能性も指摘されています。
また興味深い事に、エディアカラでは、はっきりした捕食動物の化石が1つも発見されていません。次のカンブリア紀の三葉虫化石の中には、捕食動物によって食いちぎられた様な傷跡を持つものが発見されています。しかも傷の周囲が盛り上がり、治癒の跡も見られるのです。ところがエディアカラの時代には、体が柔らかい組織でできた軟体性の生物がほとんどであるにも拘らず、生存中に傷つけられた化石は1つもないと言います。これらの事は、原生代末のエディアカラの時代には、まだ多細胞動物を捕食する肉食動物が出現していなかった事を示すものと考えられます。エディアカラの生物達は捕食される心配もなく、微生物マットで覆われた海底に横たわり、あるいは波にゆられて平和に暮らしていたのでしょう。そして、肉食の捕食者が初めて登場するのは次の古生代カンブリア紀に入ってからの事です。凶暴な捕食者のいない平和な楽園、この不思議な生物達の「エディアカラの園」は原生代の終わりと共に終焉し、顕生代はエディアカラとは全く異質な、獲物を求めて動き回る、硬い殻を持つ肉食の多細胞動物の出現によって幕が開けられるのです。
(つづく)
① 遺伝子とは何者か?
② 個体発生は系統発生を繰り返す
③ 生存闘争は進化を推進するか?
④ 生命は多様性を目指す
⑥ カンブリア爆発:捕食者の出現
⑦ 気候変動と大量絶滅

この記事が気に入ったらサポートをしてみませんか?