動物と微生物叢の相互作用における不完全な免疫が病原体の病原性を高める

メインコンテンツへスキップ
広告
インスピレーションを与える科学Cell Pressジャーナル
カレントバイオロジー
ログイン
検索...
レポート|オンライン版
動物と微生物叢の相互作用における不完全な免疫が病原体の病原性を高める
https://www.cell.com/current-biology/fulltext/S0960-9822(24)00157-X
キム・ホアン 6
ティモシー・D・リード
ケイラ・C・キング 5
脚注を表示するオープンアクセス掲載:2024年03月01日DOI:https://doi.org/10.1016/j.cub.2024.02.015
ハイライト
微生物叢による部分免疫は宿主の病原体病原性の増加を選択する
病原体の病原性と体力は進化の過程で切り離された
免疫不全宿主内での進化が病原体の多様化につながる
まとめ
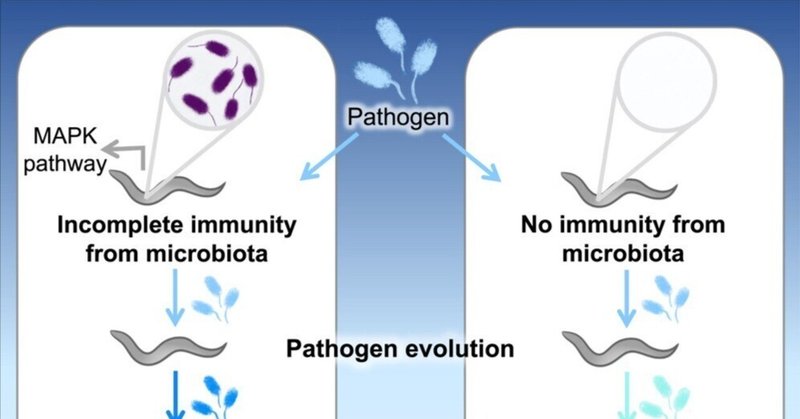
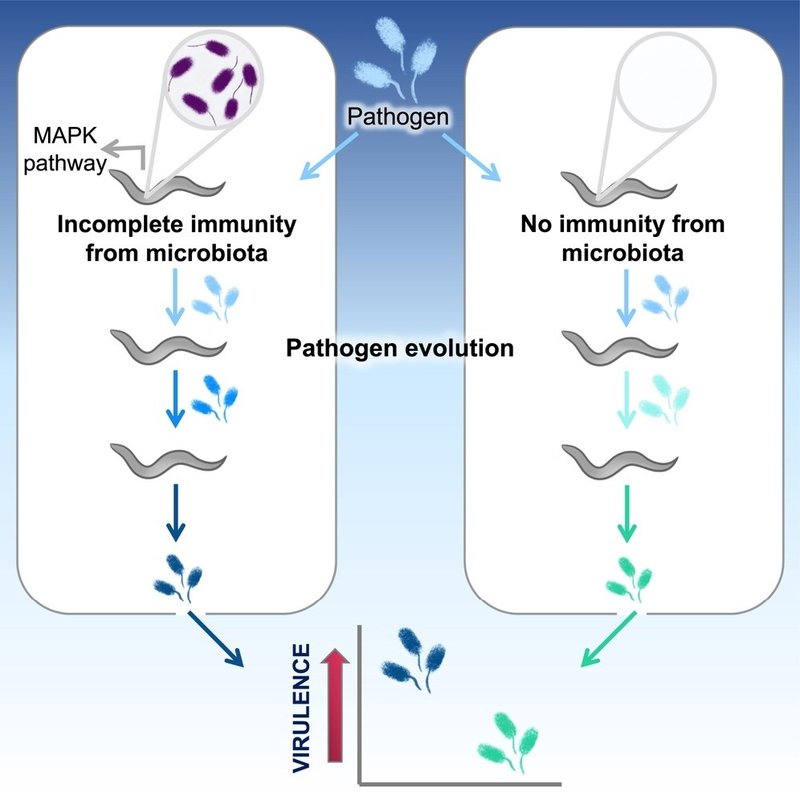
回復した宿主における不完全な免疫は、集団に再感染した際に、より病原性の強い病原体に有利に働くと予測されている1。動物をコロニー化する微生物叢は、同様に長期間持続する部分的な免疫応答を生成することができ、感染を可能にするが、疾患の重症度は低下する2。我々は、天然の微生物叢のメンバー(P. berkeleyensis)によって免疫誘導された線虫の集団を通して、実験的に継代された広範な病原体(緑膿菌)の進化の軌跡を追跡した。この細菌は、マイトジェン活性化プロテインキナーゼ(MAPK)シグナル伝達経路によって制御される遺伝子を誘導することができ、感染にもかかわらず病原体による死滅から保護するのに有効である3。宿主個体群全体において、この不完全な免疫は、プライミングされていない(すなわち、ナイーブな)、あるいは免疫不全の(MAPKオルソログをノックアウトした変異体)対照個体群で進化した病原体よりも、2倍以上死滅しやすい病原体を選択した。病原性が高いにもかかわらず、免疫増強宿主における病原体の分子進化は遅く、制約されていた。対照的に、免疫不全宿主における病原体の進化は、ゲノムの大幅な分化と病原性の減弱を特徴としていた。これらの知見は、微生物叢から誘導される不完全な宿主免疫が、新規感染症の病原性と進化のダイナミクスを形成する重要な力であることを直接的に示している。
図解抄録
図サムネイルfx1
大きな画像を見るダウンロード 高解像度画像
キーワード
病原体の進化
免疫プライミング
実験的進化
緑膿菌
線虫
結果と考察
不完全な免疫とは、病原体が再感染した場合に起こる現象で、病気の重症度や死亡率は低下する可能性が高い5、 宿主に定着している常在微生物(すなわち微生物叢)は、たとえ微生物そのものが除去されたとしても、宿主の防御的かつ長期的な免疫反応を引き起こす可能性がある6,7,8。2,10,11常在微生物と病原体との直接的な相互作用は、より病原性の低いものを選択する可能性があるが、病原体のコロニー形成が依然として起こりうる場合、免疫介在機構は逆の効果をもたらす可能性がある7,14,15。不完全な免疫は病原体に対する病原性のコストを低下させる可能性があり、この結果は感染による免疫防御の漏れがより病原性の高い病原体を好む可能性を示唆している1。
宿主微生物叢が免疫応答を介して病原体の病原性を形成しうるかどうかを直接調べるために、私たちは、広範に分布し、病気を引き起こす動物病原体(緑膿菌)を、自然な宿主-同胞間相互作用に導入して実験的に進化させた。線虫は緑膿菌に感染し、宿主の腸内に蓄積され、時間をかけて組織を破壊することにより、線虫に害を与える。P. berkeleyensisに暴露され、その後この病原体に移行した宿主は、病原体のコロニー形成と同時に常在菌を失う。しかし、P. berkeleyensisへの最初の暴露は、動植物に古くから見られる自然免疫経路であるマイトジェン活性化プロテインキナーゼ(MAPK)により制御される遺伝子を誘導するのに十分である。野生型(WT)宿主に対するP. berkeleyensisの免疫は不完全であった。P.berkeleyensisは保護された宿主にも安定した感染を形成するが、その負荷量は低かった(図1B)。P. berkeleyensisは、他の保護微生物と同様に、脅威がない場合は病原性が軽度である(図S1A)21,22。ワクチン23や脊椎動物と感染症の相互作用に関する以前の研究1とも一致するが、線虫の免疫は、宿主を病原体の伝播に制限を加えるであろう病気による死亡から保護することで、病原性のコストを低減した24。そこで我々は、微生物叢による不完全な免疫が、より病原性の強い病原体を好むかどうかを検証した。
図のサムネイルgr1
図1宿主微生物叢による不完全な免疫防御
キャプション
大きな画像を見るダウンロード 高解像度画像
我々は、P. berkeleyensisによって以前にコロニー形成された線虫個体群、あるいはナイーブな(プライミングされていない)個体群において、病原体個体群をそれぞれ独立に継代培養した(図2A)。病原体はプライム免疫反応を起こせない線虫変異体(pmk-1)でも継代した(図2A)。これらの処理は実験室適応のために宿主なしのコントロールと並行して行われた。感染時の宿主の死亡率(病原体の病原性を示す指標)と負荷量(病原体の体力を示す指標)の表現型アッセイを病原体の世代と処理に渡って行った。その後、40コロニーのプールのショットガンシーケンスを用いて、緑膿菌集団のゲノム組成の進化的変化を測定した。
図のサムネイルgr2
図2微生物叢からの不完全な免疫により、より病原性の強い病原体が選択される
キャプション
大きな画像を見るダウンロード 高解像度画像
微生物叢によって誘導された不完全免疫は、ナイーブな宿主と比較して、より病原性の高い病原体を選択した(図2B)。無脊椎動物宿主の微生物叢が、抗体を生成するワクチンと同様に病原体に影響を与えうること、そして脊椎動物において、病原体に以前さらされたことによる交差免疫が存在することから、特定のプライミング機構にかかわらず、病原性の進化における不完全免疫のより一般的な役割が示唆される。ゲノムにコードされた防御しか持たない宿主は、微生物叢を保有する免疫不全宿主と同様に、祖先の病原性レベルを維持していた。ここでは、免疫応答が弱いために、微生物叢が宿主内で長く存続できたのかもしれない。微生物叢と病原体との間の資源競争は、病原性の増加を選択すると予測されている。病原性は強くなったが、病原体は微生物叢への暴露による防御効果を克服するようには進化しなかった(図2C)。免疫プライミングは、コロニー形成が可能な強毒性病原体(例えば、WT + PM宿主に感染するWT + 保護微生物[PM]病原体)に対して、依然として害の軽減(例えば、WT - PM宿主に感染するWT + PM病原体)を提供することができる(図S1B)。
P.berkeleyensisに暴露された宿主は、ナイーブな免疫不全宿主において病原性の低下を選択したが、有意な宿主効果や相互作用効果は見られなかった(図S1D)。この結果は、プライミングされた宿主で進化する病原体の病原性がトレードオフの関係にあることを示唆している。これらの病原体は、他の進化した病原体と比較して、ナイーブな免疫コンピテントホストでは最も高い病原性を示したが、ナイーブな免疫コンピテントホストでは低い病原性を示した。進化した病原体は、微生物叢を保有する免疫不全宿主に対する影響に有意差はなかった(図S1E)。総合すると、我々の表現型から得られた知見は、微生物叢によってもたらされる生存率の向上と病原体耐性という直接的な恩恵が、最終的には宿主に極めてネガティブな影響をもたらす可能性があることを示している26。
病原体の毒性と毒性負荷は、それぞれ異なる軌跡をたどって進化した。感染時に引き起こされた宿主の死亡率と宿主あたりの細菌蓄積のレベルは、処理間で相関がなかった(図2D)。この結果は、新規病原体の病原性が実験的複製や野生個体群において独立した軌跡をたどって進化する可能性があることを示した先行研究を裏付けるものである27,28。われわれは、毒素産生や運動性など、密度に依存しない病原性因子が、免疫原性宿主の病原体において出現した高い病原性に寄与している可能性があると仮定した。病原性機構に対する選択の潜在的標的を同定するために、我々は進化した病原体集団をプール配列決定し(STAR Methods)、経時的に生じた変異を定量化した。各個体群には400-500の変異があり、ほとんどの変異は部分的に増加して個体群の50%未満であった(図S2A)。さらに処理間の一対比較から、多様な生物学的経路に関与する遺伝子における対立遺伝子頻度の違いが明らかになり(図S2B)、処理複製物には共通するユニークな変異はほとんど見られなかった(図S2CおよびS2D)。これらの結果は、宿主範囲が広い他の病原体でも見られるように、我々の実験で選択された病原性には多遺伝子的基盤があることを示唆している29,30,31。
進化した病原性の差が最も大きかった治療法(すなわち、免疫強化宿主と免疫不全宿主;図2B)の間で、集団のゲノム構成を比較した。我々は細菌べん毛機能に関与する2つの遺伝子(flgE/flgF)間に遺伝子間変異を発見した。制御領域の変異は、機能を破壊する可能性が低い。32,33 レプリケート間の変異頻度は、感染宿主の死亡率と正の相関があった(図S3AおよびS3B)。鞭毛は病原性因子34,35であり、運動性に必要であるため、進化した集団の遊泳能力を比較した(STAR Methods; 図2、挿入図)。病原体の運動性は、これらの極端な処理間で有意に異なったが(図2E)、すべての処理での差はわずかであった(図S3C)。各病原体集団のわずかな割合(30%未満)しかflgE/flgF変異を持たなかったことから(図S3A)、flgE/flgF変異が病原性の唯一の要因ではないことが示唆された。この変異を持つ細胞の部分集団は、他の変異を持つ細胞と相互作用している可能性がある。緑膿菌は、免疫のある宿主と免疫のない宿主に適応するために異なる遺伝的経路を利用したが、両グループは同じようなフィットネスレベルに収束した39,40。
微生物叢によって誘導される宿主の免疫応答の強さは、新規病原体のゲノム進化を形作る可能性がある。免疫不全宿主に見られるような弱い淘汰のもとで病原体が複製されると41、突然変異が蓄積しやすくなり、ゲノムの多様化が進む。このようなゲノム進化の急速な変化は、人獣共通感染症の原因となる細菌性病原体42,43やウイルス性病原体44で示されている。
個々の病原体のコロニーと祖先との関係を評価するために、点突然変異に基づく系統樹を構築した(図3)。個々のコロニーで同定された変異のほとんどは、プールされたサンプルでも固定されていた(図S4A-S4E)。免疫不全宿主で進化する病原体は、祖先から大きく分岐していた(個々のコロニーあたり5.57±0.80変異;図3)。また、これらのコロニーは、in vitroで進化したコロニーと同様に祖先からの距離を共有し(図3およびS4F)、さらに、同様の病原性レベルに収束した(図2B)。acoA(Acetoin dehydrogenase E1 component α-subunit)遺伝子は、免疫のない宿主で進化した病原体では、免疫のある宿主で進化した病原体に比べて変異が多く、非同義および小さなインデルの割合が高い(図S4G-S4K)。KEGG(Kyoto Encyclopedia of Genes and Genomes)パスウェイ解析により、この遺伝子が多様な環境における微生物代謝、代謝経路、二次代謝産物の生合成に関与していることが明らかになった46。fmtと同様に、acoAの変異はこれらの病原体が示す病原性の低下に何らかの役割を果たしている可能性がある。これらの結果は、弱い選択によって獲得された変異が病原性を低下させ、遺伝的多様性を増加させることを示している。同様の結果は、免疫系に欠陥のある宿主に感染する病原体でも見つかっており42,47,48、そこでは病原性の低い病原体はより病原性の高い病原体に対してより有利に競争できる可能性がある49。高い病原性を強く選択したにもかかわらず、不完全な免疫は最終的に病原体の進化を分子レベルで制限した。
図のサムネイルgr3
図3微生物叢からの不完全な免疫が病原体の分子進化を抑制する
キャプション
大きな画像を見るダウンロード 高解像度画像
宿主の免疫応答が病原体集団間の分岐の度合いを変化させた。宿主の防御が弱いと(すなわち、選択が弱いと)、防御が強い宿主と比較して、集団間で病原体の遺伝的分岐が大きくなる可能性がある50,51,52,53。41 宿主の防御が病原体集団の分岐にどのような影響を与えたかを調べるために、各処理内の複製集団間で各SNPについてペアワイズFSTを計算した。免疫原性宿主で進化した病原体は、ゲノムコード化された防御によってのみ保護された宿主で進化した病原体に比べて、有意なFST遺伝子座が少なかった(図4A)。微生物叢が存在しないことは、処理全体にわたって有意なFST遺伝子座の増加に寄与したが、この効果は2つのWT宿主処理間の違いによってもたらされた可能性が高い。宿主効果は見られず、この研究では検証されなかった他の選択的な力(例えば、免疫不全宿主における微生物叢と病原体の間の資源競争)による可能性がある。すべての処理で、細菌分泌系と2成分系に関与する遺伝子に分化が認められた。これらの結果は、宿主微生物叢によって生成される不完全な免疫は、非刺激処理と比較して、複製集団間の遺伝的分化を制限したことを示している。
図サムネイルgr4
図4 微生物叢が誘導する宿主防御が病原体の進化経路を変える
キャプション
大きな画像を見るダウンロード 高解像度画像
また、実験の中間点(すなわち、継代7[P7])と終点において、祖先病原体と進化した集団との間のFSTを計算することにより、集団全体の遺伝的組成の時間的シフトを評価した。中間点では、免疫のない宿主で進化した病原体の方が、免疫のある宿主で進化した病原体よりも有意なFST遺伝子座を持っていた(図4B、「祖先対P7」)。微生物叢が存在しないと、祖先とP7との間で有意な遺伝子座の数が増加し(p = 0.011)、特に免疫不全宿主では有意な遺伝子座の数が増加した(p = 0.006)。P7と継代14(P14)の間ではより少ない差異しか検出されなかった。ゲノムにコードされた防御機能しか持たない宿主で進化した病原体集団は、免疫のある宿主で進化したもの(p = 0.034)および免疫のない宿主で進化したもの(p = 0.045)よりも、経時的な差が大きかった。実験が終わるころには、有意なFST遺伝子座の数という点では、もはや処理に差はなくなっていた(図4B、「祖先対P14」)。進化の初期には、常在細菌叢が存在しないため、祖先と進化した病原体との間に遺伝的差異が生じたが、最終的にはすべての集団が同様の変化率を示した。これらの結果を総合すると、病原体の出現初期における進化を形成するダイナミクスは、適応の時期を過ぎると異なるものになる可能性があることが示唆される43,45。
宿主微生物叢は、有害な感染から宿主を守る上で、生命の木全体にわたって重要な役割を果たすことができる。11,54,55 しかし、進化の過程で、宿主微生物叢によって誘導される不完全な免疫は、リーキーワクチン23,56や過去の感染1による進化予測と同様に、強毒性病原体を好むように作用する可能性があることがわかった。逆に、免疫不全に陥った宿主は、病原体が変異を蓄積しやすい環境となり、ゲノムの分解や宿主の制限につながる可能性がある42。したがって、宿主微生物叢と免疫の相互作用は、新興感染症の進化を形成する淘汰の主要な原因となっている可能性がある。プロバイオティック微生物の利用は、農業や野生動物の生態系に浸透しつつある57,58。我々は、病原体の進化にもかかわらず、これらの微生物治療薬の有効性が維持される可能性があることを発見した。しかしながら、より病原性の強い病原体の亜種の拡散を促進する可能性がある場合には、将来のリスクと宿主個体への直接的な利益とのバランスを考慮した上で、適切な予防措置を講じる必要がある。
STAR★メソッド
主要リソース表
試薬またはリソースソースの識別子
化学物質、ペプチド、組み換えタンパク質
Lysogeny broth (LB) 培地 オックスフォード大学生化学部の培地キッチンで調製 N/A
M9培地 オックスフォード大学生化学科、培地キッチンで調製 N/A
線虫増殖培地(NGM)寒天 オックスフォード大学生化学科メディアキッチンで調製 N/A
Triton X-100 Sigma-Aldrich CAS番号:9036-19-5
重要な市販アッセイ
DNeasy Blood and Tissue Kit Qiagen カタログ番号。69504
寄託データ
祖先および進化した病原体集団の全ゲノムリシークエンスデータ National Center for Biotechnology Information (NCBI) BioProject: PRJNA998467
Phenotypic and processed genomic data Mendeley Data メンデレーデータ: https://doi.org/10.17632/xz9t9gjtw6.1
実験モデル 生物/株
生物 線虫N2株 ブリストル線虫遺伝学センターN2
生物: 線虫pmk-1株 (M03F8.4(op497), pmk-1(km25)) Jonathan Hodgkin (オックスフォード大学) Km25
生物 大腸菌OP50 ケノラブディティス・ジェネティクスセンター OP50
生物 緑膿菌 PA14-GFP Kevin Foster(オックスフォード大学) PA14
生物 Pseudomonas berkeleyensis MSPm1 Michael Shapira(カリフォルニア大学バークレー校) MSPm1
ソフトウェアとアルゴリズム
R バージョン 4.2.0 https://www.r-project.org 該当なし
Breseq https://barricklab.org/twiki/pub/Lab/ToolsBacterialGenomeResequencing/documentation/ N/A
Popoolation2 https://sourceforge.net/p/popoolation2/wiki/Home/ N/A
PHYLIP https://phylipweb.github.io/phylip/ 該当なし
新しいタブで表を開く
リソースの有無
連絡先
リソースや資料に関するさらなる情報やリクエストは、リードコンタクトであるKim Hoang (kim.hoang@emory.edu)までお願いします。
資料の入手可能性
進化した個体群は、Kim Hoangからリクエストにより入手可能です。
データおよびコードの入手可能性
生配列はBioProject:PRJNA998467としてNCBI Sequence Read Archiveに寄託された。表現型データはMendeley Data:https://doi.org/10.17632/xz9t9gjtw6.1。
本論文の解析に使用したRコードは、リクエストに応じて入手可能である。
本論文で報告されたデータを再解析するために必要な追加情報は、要請があれば主任連絡先から入手可能である。
実験モデルと被験者の詳細
N2およびpmk-1線虫は-80℃で保存したストックから作製し、線虫増殖培地(NGM)プレート上で大腸菌OP50とともに20℃で維持した。Pseudomonas berkeleyensis MSPm1、緑膿菌 PA14-GFP、および大腸菌 OP50 は、-80℃で保存したストックから樹立し、溶菌ブロス(LB)寒天培地プレート上で 30℃で一晩培養した。シュードモナス・バークレーエンシスおよび大腸菌は、実験的進化および各アッセイごとに凍結ストックから復活させた。線虫のストック個体群は、実験的進化とアッセイの間、-80℃から定期的に復活させた。
方法の詳細
祖先の緑膿菌を用いた生存率とCFUアッセイ
宿主の生存
細菌を準備するため、P. berkeleyensis、大腸菌、緑膿菌のランダムなコロニーを1個ずつ、LB中、30℃の振盪培養器で一晩培養した。その後、9cmのNGMプレートに菌を播種し、30℃で1日培養した。N2およびpmk-1線虫の卵を集め、表面殺菌し、標準的な次亜塩素酸ナトリウムのプロトコールに従って年齢同調させた63。孵化後、約200匹のL1幼虫をNGM上のP. berkeleyensisまたはE. coliの芝生にスポットした。これらの線虫を 20℃で 2 日間培養した。その後、L4/幼成虫をNGM上の緑膿菌の芝生に移し、20℃で保温した。3日後、プラチナピックで線虫を突いて移動の兆候を調べ、生きている線虫の数を決定した。
病原体CFU
上記の手順で線虫を3日間感染させた後、VegaとGore64のプロトコルを修正して、感染した線虫の病原体量を測定した。簡単に説明すると、1集団あたり10匹の線虫を採取し、0.01% Triton X-100(M9-T)を含む冷M9緩衝液で2回洗浄した後、氷上で約30分間冷やし、蠕動を停止させた。その後、線虫/M9混合液の最終濃度が0.3%になるように十分な量の冷漂白剤を加えた。ざっと混合した後、混合液を氷上で10分間静置し、冷M9-Tを加えて漂白を止めた。線虫を冷M9-Tでもう一度洗浄し、上清をプレーティングして漂白の効率を確認した。解剖スコープの下で、約100ulのM9-Tにジルコニウムビーズを入れた別のチューブに10個体をピペットで入れた。サンプルをTissueLyserのビーズビーターで27 1/sで2分間振盪した。短時間の遠心分離後、連続希釈したホモジネートを9cmのLB寒天プレートに広げ、30℃で培養した。日後にコロニー形成単位数を定量した。
宿主の繁殖力
N2 または pmk-1 線虫を P. berkeleyensis または E. coli で飼育した。P.berkeleyensisまたはE. coliを播種した9cm NGMプレート上でL1をL4/若成虫まで飼育した(20℃で約2日間)。3日後、緑膿菌に感染した線虫の宿主死亡率を測定したのと同じ日に、顕微鏡下で幼虫の数を数えた。
実験的進化
緑膿菌PA14-GFPを5つの処理(図2A)で継代した:4つの宿主処理と1つの宿主なし処理。まず、PA14-GFPのコロニー1個をLBブロス中で一晩増殖させ、線虫増殖培地65上にまいた後、30℃で1日培養した。約1000匹の線虫をそれぞれの飼育プレート(後述)から緑膿菌プレートに移し、20℃で培養した。線虫は1日後に各プレートから洗浄し、M9バッファーで3回すすいだ。M9/線虫混合液の10%をビーズビーターで破砕し、ホモジネートをLBプレートにプレーティングした。一晩培養した後、100コロニーをブロスに摘み取り、次の継代を開始した。各処理は14継代にわたり、5反復の飼育と緑膿菌プレートで構成された。
実験期間中、線虫は進化的に静止させた(進化させない)。N2およびpmk-1個体群は、祖先の緑膿菌のセクションの生存およびCFUアッセイで説明したように飼育した。L4/若齢成虫は上記のように緑膿菌プレートに移した。それぞれの継代では、実験期間中、宿主系統におけるde novo突然変異の蓄積を抑えるため、-80℃から定期的に復活させたストック線虫集団から卵を集めた。
進化した緑膿菌を用いた死亡率とCFUアッセイ
進化した個体群の死亡率とCFUアッセイは、祖先緑膿菌の場合と同様のプロトコールに従った(祖先緑膿菌の生存率とCFUアッセイの項)。アッセイは3連で行った。図2Bと2Dについては、OP50で飼育したN2線虫を感染させた。図 2C と S1B では、P. berkeleyensis で飼育した N2 線虫を感染させた。図S1CとS1Dについては、それぞれ大腸菌またはP. berkeleyensisで飼育したpmk-1線虫を感染させ、これらの宿主では死亡率が高いため、3日後ではなく2日後に死亡率を定量した。
遊泳運動性
祖先と進化した緑膿菌の運動性を測定するために、Haら66のプロトコールに従って遊泳運動性プレートを接種した。プレートは30℃で1日培養した。これはNGMプレートが線虫を病原体上に置く前に培養されていた温度であったためである。この日の細菌増殖の直径を初期直径とした。その後、宿主の死亡率を測定するために、感染タイムラインから3日間、20℃でプレートを培養し、最終直径を測定した。最終直径から初期直径を引いて遊泳直径の変化を求めた。
DNA抽出と配列決定
プールサンプルの場合、各レプリケート集団について40個のコロニーをLBブロスで一晩別々に増殖させ、各コロニーのOD600を標準化してから1つのチューブにプールしてDNA抽出を行った。単一コロニーサンプルの場合は、個々のコロニーをLBブロスで一晩別々に増殖させた後、DNA抽出を行った。DNeasy Blood and Tissue Kit(Qiagen)を用いて、製造元の指示に従ってゲノムDNAを抽出した。サンプルライブラリーはIllumina DNA Prep kitを用いて調製し、Illumina NextSeq 2000でシーケンスした。配列品質はFastQC(https://www.bioinformatics.babraham.ac.uk/projects/fastqc/)を用いて評価し、配列はfastPを用いてトリミングした67。各宿主処理の5つの複製集団すべてと、宿主処理なしの3つのランダム集団の配列を決定した。単一コロニーサンプルについては、WT+PMおよびIC-PM処理の各集団から無作為に3個体、その他の処理の各集団から無作為に1個体のコロニーの塩基配列を決定した。
祖先のPA14-GFP個体コロニーは、ハイブリッドアセンブリのためにイルミナに加えてOxford Nanopore Technologies(ONT)を用いて配列決定した。品質管理およびアダプタートリミングは、それぞれイルミナおよびONTシーケンス用のデフォルトパラメーターを用いて、bcl2fastq68およびporechop69で行った。イルミナおよびONTリードのハイブリッドアセンブリはUnicyclerで行い70、得られたアセンブリはBaktaアノテーションパイプラインを用いてアノテーションした71。各プールサンプルのカバレッジは200倍以上、各シングルコロニーサンプルのカバレッジは60倍以上であった。
定量化と統計解析
表現型データおよび処理されたゲノムデータの統計解析はすべてRバージョン4.2.0で行った。有意閾値はP < 0.05とした。図中のエラーバーは標準誤差を表す。各アッセイのサンプルサイズは図の凡例に示した。
祖先病原体データの解析
祖先病原体に感染した宿主の死亡率データは、二項分布による一般化線形混合モードで解析し、Tukey多重比較検定で一対の差を決定した。祖先病原体のCFUデータはt検定で解析し、宿主の繁殖力データはANOVAで解析した。
進化した病原体データの解析
P. berkeleyensisで飼育したpmk-1の死亡率データは線形混合モデルを用いて解析した。P. berkeleyensisで飼育したN2の病原体CFUデータは、正規性の仮定を満たすように平方根変換し、線形混合モデルを用いて解析した。他のすべての宿主の死亡率および残りの CFU データは、一般化線形混合モデル(それぞれ二項分布またはポアソン分布)を用いて解析し、Tukey 多重比較検定を用いて一対の差異を判定した。運動性データは線形混合モデルを用いて解析した。
プールサンプルの解析
Breseq pipeline polymorphism modeを用い、デフォルトパラメーターを用いて、祖先を参照としてバリアントをコールした74。flgE/flgFおよびfmt変異の頻度が死亡率と相関しているかどうかは、スピアマンの順位相関を用いて検定した。
治療法間の対立遺伝子頻度の差を決定するために、各治療法の全レプリケート集団のリードをプールした。その後、Popoolation2パイプライン75を用いて正確な対立遺伝子頻度差を計算し、フィッシャーの正確検定を用いて有意性を推定した。コーディング領域で見つかった有意な遺伝子座については、Database for Annotation, Visualization and Integrated Discovery (DAVID)76ツールを用いて、各遺伝子をKEGGパスウェイにマッピングした77。
各治療におけるSNPごとのFSTを計算するために、Popoolation2パイプラインを治療内の複製集団のすべてのペアワイズの組み合わせに使用した。時点間のSNPごとのFSTを計算するために、Popoolation2パイプラインを使用して、各集団を7番目の継代と祖先、14番目と7番目の継代、14番目と祖先の間で比較した。両分析とも、フィッシャーの正確検定を用いて有意性を推定した。その後、ボンフェローニ補正後に有意な遺伝子座の数を数えた。各処理区間ではカイ二乗検定を用いて集団間の遺伝子座数を比較し、各時点間では線形モデルまたはポアソン分布付き一般化線形モデルを用い、Tukey多重比較検定を用いて一対の差を決定した。
個々のコロニーサンプルの解析
デフォルトのパラメータでbreseqパイプラインを使用して、祖先を参照としてバリアントをコールした。次に、ape79のcophenetic.phylo関数を用いて、祖先と各コロニー間の一対距離を計算した。データは線形混合モデルを用いて分析した。
謝辞
Dana HawleyとLevi Morranには原稿に対するフィードバックを、King、Read、Morranの各研究室のメンバーには洞察に満ちた議論をいただいた。また、Steve Diggleには緑膿菌について、Julia Kreinerには集団ゲノムについて、Jelly Vanderwoudeにはハイパームテーターについてアドバイスをいただいた。また、SeqCenter (seqcenter.com)には、ハイスループットシーケンスデータの作成と、緑膿菌の祖先リファレンスゲノムのアセンブルにご協力いただいた。K.L.H.は、NSFのPostdoctoral Research Fellowship in Biology (1907076)およびAmerican Association of University WomenのResearch Publication Grant in Engineering, Medicine, and Scienceの資金援助を受けた。K.C.K.は、European Research Council Starting Grant(COEVOPRO 802242)およびNSERC Canada Excellence Research Chairの助成を受けた。
著者貢献
K.L.H.とK.C.K.は本研究の構想および設計を行った。K.L.H.はT.D.R.およびK.C.K.の指導の下、データ収集とデータ解析を行った。K.L.H.とK.C.K.は論文の草稿を作成し、全著者による批判的な修正を加えた。
利害関係
著者らは、競合する利益はないと宣言している。
補足情報
pdfをダウンロード (2.04 MB)
pdfファイルのヘルプ
ドキュメントS1. 図S1-S4および表S1、S2
参考文献
フレミング・デイビーズ A.E.
ウィリアムズ P.D.
ドントA.A.
ドブソン A.P.
ホーチャッカW.M.
レオン A.E.
レイ D.H.
オスナス E.E.
ホーリーD.M.
宿主免疫の不完全さが、新興病原体における病原性の進化を促進する。
Science. 2018; 359: 1030-1033
論文で見る
スコープス (40)
PubMed
クロス
グーグル奨学生
ホアン K.L.
キング K.C.
動物における共生生物介在性免疫プライミングの進化論的考察。
微生物学。2022; 168: 1-11
論文で見る
スコパス(2)
クロスリファレンス
グーグル奨学生
モンタルボ・カッツ S.
ホアン・H.
アペル M.D.
バーグ M.
シャピラ M.
線虫のp38依存性感染抵抗性を高める土壌細菌との会合。
Infect. Immun. 2013; 81: 514-520
論文で見る
スコープス (94)
パブコメ
クロス
グーグル奨学生
ミルチノビッチB.
カーツJ.
無脊椎動物における免疫記憶。
Semin. Immunol. 2016; 28: 328-342
論文で見る
スコープス (192)
PubMed
クロス
グーグル奨学生
レオン A.E.
ホーリーD.M.
自然界の鳴禽類の宿主における病原体のプライミングに対する宿主の反応。
Ecohealth. 2017; 14: 793-804
論文で見る
スコパス (15)
クロスリファレンス
グーグル奨学生
キム Y.
Mylonakis E.
プロバイオティクス細菌であるラクトバチルス・アシドフィルス ncfm株による線虫の免疫コンディショニングはグラム陽性免疫応答を増強する。
Infect. Immun. 2012; 80: 2500-2508
論文で見る
日本
PubMed
クロス
グーグル奨学生
クウォン W.K.
マンセニド A.L.
モラン N.A.
ミツバチの自生腸内細菌叢による免疫系刺激。
R. Soc. Open Sci. 2017; 4170003
論文で見る
スコープス (237)
PubMed
クロスフィルム
グーグル奨学生
クラーク T.B.
デイビス K.M.
リセンコ E.S.
Zhou A.Y.
Yu Y.
ワイザーJ.N.
Nod1による微生物叢からのペプチドグリカンの認識は、全身の自然免疫を増強する。
Nat. Med. 2010; 16: 228-231
論文で見る
スコパス (889)
PubMed
クロス
グーグル奨学生
セロッセ M.A.
ベシスA.
ポゾM.J.
微生物による動植物免疫のプライミング:発生シグナルとしての共生生物。
Trends Microbiol. 2014; 22: 607-613
論文で見る
スコープス (75)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ガブリエリP.
カッチャ S.
ヴァロット・ボッカッツィ I.
アルノルディ I.
バルビエリ G.
コマンダトーレ F.
エピス S.
蚊の三部作:微生物相、免疫、病原体、そして疾病伝播の制御に対するそれらの意味合い。
フロント。Microbiol. 2021; 12630438
論文で見る
スコパス (39)
クロスリファレンス
グーグル奨学生
ホスラビ A.
マズマニアン S.K.
微生物感染症の危険因子としての腸内マイクロバイオームの破壊。
Curr. Opin. Microbiol. 2013; 16: 221-227
論文で見る
スコープス (148)
パブコメ
クロス
グーグル奨学生
フォード S.A.
カオ D.
ウィリアムズ D.
キング K.C.
微生物を介した宿主防御が病原体の病原性低下の進化を促進する。
Nat. Commun. 2016; 713430
論文で見る
スコパス (47)
クロスリファレンス
グーグル奨学生
ネルソン P.
メイG.
防御的共生と病原性の進化。
Am. 2020; 196: 333-343
論文で見る
スコパス (6)
クロスリファレンス
グーグル奨学生
ホラックR.D.
レナード S.P.
モラン N.A.
共生生物はミツバチの宿主自然免疫を形成する。
Proc. Biol. サイエンス 2020; 28720201184
論文で見る
Google Scholar
コービー-ハリス V.
スナイダーL.
ミードールC.A.D.
ナルドR.
モット B.
アンダーソン K.E.
Parasaccharibacter apium, gen. Nov., sp.はミツバチ(Hymenoptera: Apidae)のノセマに対する抵抗性を改善する。
J. Econ. Entomol. 2016; 109: 537-543
論文で見る
スコープス (0)
PubMed
クロス
グーグル奨学生
イラゾキ J.E.
トロイメルE.R.
ファインバウムR.L.
ルハチャックL.G.
セザイヤン・B.O.
アウズベル F.M.
緑膿菌と黄色ブドウ球菌による線虫感染における異なる病原性と宿主応答。
PLoS Pathog. 2010; 6e1000982
論文で見る
(254件)
PubMed
クロス
グーグル奨学生
ダークセン P.
マーシュ S.A.
ブレイカーI.
ハイトランドN.
ワグナー S.
ナカド R.
マダー S.
ピーターセン C.
コワリク V.
ローゼンスティールP.
他
線虫Caenorhabditis elegansのネイティブマイクロバイオーム:新しい宿主マイクロバイオームモデルへのゲートウェイ。
BMC Biol.
論文で見る
スコープス (225)
PubMed
クロスフィルム
グーグル奨学生
ディルクセン P.
アシエA.
ツィンマーマンJ.
チャン F.
ティーチェ A.M.
マーシュ S.A.
フェリックス M.A.
シャピラ M.
カレタ C.
シューレンブルグH.
他。
CeMbio - 線虫マイクロバイオームリソース。
G3 (Bethesda). 2020; 10: 3025-3039
論文で見る
(61件)
PubMed
クロス
グーグル奨学生
グレース A.
サフ R.
オーウェン D.R.
デニス V.A.
緑膿菌基準株PAO1およびPA14:ゲノム、表現型、および治療法のレビュー。
Front. Microbiol. 2022; 131023523
論文で見る
スコパス (14)
クロスリファレンス
グーグル奨学生
エンゲルマン I.
プジョル N.
線虫における自然免疫。
Adv. Exp. Med. 2010; 708: 105-121
論文で見る
PubMed
クロスフィルム
グーグル奨学生
Vorburger C.
ガネサナンダモアティP.
Kwiatkowski M.
アブラムシの寄生虫に対する共生生物抵抗性の構成コストと誘導コストの比較。
Ecol. 2013; 3: 706-713
論文で見る
日本農芸化学会誌
PubMed
クロス
グーグル奨学生
Łukasik P.
グオ H.
ヴァン・アッシュ M.
フェラーリ J.
ゴッドフレイ H.C.J.
リケッチアとスピロプラズマの共生によってもたらされる真菌病原体に対する防御は、複数の宿主の遺伝子型と種で発現し、別の共生生物との同時感染には影響されない。
J. 進化生物学 2013; 26: 2654-2661
論文で見る
スコパス (96)
PubMed
クロス
グーグル奨学生
リード A.F.
ベイジェント S.J.
パワーズC.
クゴサナ L.B.
ブラックウェル L.
スミス L.P.
ケネディ D.A.
ウォークデン・ブラウン S.W.
ネール V.K.
不完全なワクチン接種は、強毒性病原体の伝播を促進する可能性がある。
PLoS Biol.
論文で見る
スコープス(259)
クロスリファレンス
グーグル奨学生
パイク V.L.
スティーブンス E.J.
グリフィン A.S.
キング K.C.
生産者病原体と非生産者病原体の宿主内および宿主間動態。
寄生虫学。2023; 150: 805-812
論文で見る
スコパス (0)
クロスリファレンス
グーグル奨学生
フォルブルガー C.
パールマン S.J.
宿主と寄生虫の共進化における防御的共生生物の役割。
Biol. Rev. Camb. Philos. Soc. 2018; 93: 1747-1764
論文で見る
スコープス (0)
PubMed
クロス
グーグル奨学生
スミス C.A.
アシュビーB.
寛容をもたらす防御共生生物と寄生虫の病原性の進化。
Evol. 2023; 7: 262-272
論文で見る
クロスリファレンス
グーグル奨学生
タルディ L.
ジロドー M.
ヒル G.E.
マクグロー K.J.
ボンノー C.
新興細菌病原体における病原性と複製速度の対照的進化。
Proc. Natl. Acad. Sci. USA. 2019; 116: 16927-16932
論文で見る
スコープス (21)
Crossref
グーグル奨学生
エクロス A.K.E.
ゲルト M.
スティーブンス E.J.
フォード S.A.
キング K.C.
宿主の遺伝子型と遺伝的多様性が新規細菌感染の進化を形成する。
2021年; 15: 2146-2157
論文で見る
スコープス (13)
クロスリファレンス
グーグル奨学生
チェン・エイチ
ブライト・R.A.
スバラオ K.
スミス C.
コックス N.J.
カッツ J.M.
松岡洋一
1997年香港型H5N1インフルエンザウイルスのマウスにおける病原性に関与する多遺伝子病原性因子。
ウイルス研究 2007; 128: 159-163
論文で見る
スコープス (112)
PubMed
クロスフィルム
グーグル奨学生
ル・クレックW.
シュバリエF.D.
マクデュー-ホワイトM.
メノン V.
アーリア G.A.
アンダーソン T.J.C.
住血吸虫の感染ステージ産生と病原性の遺伝的構造。
Virulence. 2021; 12: 1508-1526
論文で見る
スコパス (6)
クロスリファレンス
グーグル奨学生
ケイシス C.
シーG.
ソルティスN.
グウィナー R.
コーウェン J.
アトウェル S.
クライベンシュタイン D.J.
定量的相互作用:植物界におけるボトリティス・シネレアの発病結果。
G3(ベセスダ)。2021; 11jkab175
論文で見る
スコープス (10)
クロスリファレンス
グーグル奨学生
シャープ C.
フォスターK.R.
ジェンナ・V.
ヴィドシッチ P.
ソマロフトゥS.
パイル A.M.
デ・ヴィボ M.
マーシャ M.
著者訂正: スプライシングの第一段階と第二段階の間のグループIIイントロンのダイナミクスの可視化。
Nat. Commun. 2022; 131
論文で見る
グーグル奨学生
ロッセ・Y.
ウォルフソン E.B.
ホームズA.
ガリー D.L.
ホールデン N.J.
細菌鞭毛:ねじり、くっつき、あるいは王国を越えてかわす。
PLoS Pathog. 2015; 11e1004483
論文で見る
スコープス(118)
クロスリファレンス
グーグル奨学生
ファインバウム R.L.
アーバック J.M.
リベラティ N.T.
ジョノビッチ S.
アドニツィオ A.
カルブニスA.R.
アウズベル F.M.
線虫感染モデルを用いた緑膿菌病原性関連遺伝子のゲノムワイド同定。
PLoS Pathog. 2012; 8e1002813
論文で見る
スコパス (123)
クロスリファレンス
グーグル奨学生
Duan Q.
Zhou M.
Zhu L.
Zhu G.
鞭毛と細菌の病原性。
J. Basic Microbiol. 2013; 53: 1-8
論文で見る
スコープス (214)
パブコメ
クロス
グーグル奨学生
マルコV.A.
キルムリーS.L.N.
マクニールL.T.
バローズ L.L.
緑膿菌IV型マイナーピリンとPilY1は、FimS-AlgR活性を調節することにより病原性を制御する。
PLoS Pathog. 2018; 14e1007074
論文で見る
スコパス (39)
クロスレビュー
グーグル奨学生
ルイス・ベドヤ T.
ワン・P.W.
デスヴォー D.
ガットマン D.S.
分泌型病原体エフェクターの集団作用を介した協調的病原性。
Nat. Microbiol. 2023; 8: 640-650
論文で見る
スコパス (9)
クロスリファレンス
グーグル奨学生
ギヨン J.M.
メクラムY.
シュミッターJ.M.
ブランケ S.
ファヤットG.
Met-tRNA(f)/(Met)ホルミル転移酵素遺伝子の破壊は大腸菌の増殖を著しく阻害する。
J. Bacteriol. 1992; 174: 4294-4301
論文で見る
スコープス (0)
PubMed
クロス
グーグル奨学生
ベイリー S.F.
ロドリゲ N.
Kassen R.
並列進化の確率に及ぼす選択環境の影響。
Mol. Biol. Evol. 2015; 32: 1436-1448
論文で見る
スコープス (81)
PubMed
クロス
グーグル奨学生
フリッケルJ.
フォイルナーP.G.D.
カラコックE.
ベックス L.
宿主-ウイルス系において、集団サイズの変化と選択が並行進化のパターンを駆動する。
Nat. Commun. 2018; 91706
論文で見る
スコープス (22)
クロスリファレンス
グーグル奨学生
バローゾ=バティスタ J.
デメンゲオ J.
ゴルド I.
適応免疫は、常在腸内細菌の進化的変化のペースと予測可能性を高める。
Nat. Commun. 2015; 68945
論文で見る
スコープス (61)
クロスリファレンス
グーグル奨学生
クレム E.J.
グクラニア-クロツァスE.
ハドフィールドJ.
フォーブスターJ.L.
ハリス S.R.
ヘイル C.
ヒース J.N.
ワイルマン T.
クレア S.
ケイン L.
他。
免疫不全宿主における急速な進化による宿主適応型サルモネラ・エンテリティディスの出現。
Nat. Microbiol. 2016; 1: 1-6
論文で見る
スコパス (67)
クロスリファレンス
グーグル奨学生
ローネイ A.
ウー C.J.
デュラント・チェンA.
ユン J.H.
キル P.P.
デッカー J.P.
免疫不全ヒト宿主における新興人獣共通感染症細菌病原体のin vivo進化。
Nat. Commun. 2021; 124495
論文で見る
スコパス (5)
クロスリファレンス
グーグル奨学生
ケンプ S.A.
コリアー D.A.
ダティール R.P.
フェレイラ I.A.T.M.
ゲイド・S.
ジャーウン A.
ホスミロ M.
リーズ=スピア C.
ムルコチョバ P.
ラムI.U.
他。
慢性感染症治療中のSARS-CoV-2の進化。
Nature. 2021; 592: 277-282
論文で見る
スコープス (587)
PubMed
クロス
グーグル奨学生
デイ T.
ケネディ D.A.
リードA.F.
ガンドン S.
ワクチン接種キャンペーン中の病原体の進化。
PLoS Biol.
論文で見る
スコープス (6)
クロスリファレンス
グーグル奨学生
ウィンザーG.L.
グリフィス E.J.
ロー R.
ディロン B.K.
シェイ J.A.
ブリンクマン F.S.
Pseudomonasゲノムデータベースの数千のPseudomonasゲノムを比較するためのアノテーションと機能の強化。
Nucleic Acids Res.
論文で見る
日本学術振興会特別研究員
PubMed
Crossref
グーグル奨学生
モンジュリ V.
レキームS.
クサタナスA.
ゴーソン V.
ブラン H.
ニグ J.
キンタナ=ムルチ L.
エレナ S.F.
サレ M.C.
自然免疫経路が相乗的に働き、キイロショウジョウバエのRNAウイルス進化を抑制している。
Nat. Ecol. 2022; 6: 565-578
論文で見る
スコパス (7)
PubMed
クロス
グーグル奨学生
ヤンセンG.
クラムネルL.L.
ジルベールF.
モア T.
プフェフェルコルンR.
テーナート R.
ローゼンスティール P.
Schulenburg H.
病原性から通性への進化的移行: グローバルレギュレーター変異は病原性の減衰を通じてフィットネスの向上を媒介する。
Mol. Biol. Evol. 2015; 32: 2883-2896
論文で見る
スコパス (36)
PubMed
クロス
グーグル奨学生
Råberg L.
デ・ルードJ.C.
ベル A.S.
スタムー P.
グレイ D.
リード A.F.
遺伝的に多様なマラリア感染における免疫介在性の見かけ上の競合の役割。
Am. Nat. 2006; 168: 41-53
論文で見る
スコパス (0)
PubMed
クロス
グーグル奨学生
クルックシャンク T.
ウェイド M.J.
ショウジョウバエにおける遺伝子発現パターンが配列変異と分岐を形成する。
Evol. Evol. Dev. 2008; 10: 583-590
論文で見る
スコープス (70)
PubMed
クロス
グーグル奨学生
ルネマルクA.
ブライデゴーM.
スヴェンソン E.I.
緩やかな捕食は島嶼個体群間の表現型の分岐を促進するか?
J. Evol. Biol.
論文で見る
スコパス (26)
クロスリファレンス
グーグル奨学生
マクファーソン A.
ニュイスマー S.L.
常在遺伝子変異からの並列遺伝進化の確率。
J. Evol. Biol. 2017; 30: 326-337
論文で見る
スコープス (33)
PubMed
クロス
グーグル奨学生
スクリブナー M.R.
サントス-ロペスA.
マーシャルC.W.
デイトリックC.
クーパーV.S.
ホーガン D.A.
種や環境を超えたトブラマイシン耐性の並列進化。
mBio. 2020; 11 (e00932-e00920)
論文で見る
(37件)
PubMed
クロス
グーグル奨学生
キング K.C.
防御共生生物
Curr. Biol. 2019; 29: R78-R80
論文で見る
スコープス (19)
PubMed
要旨
全文
全文PDF
グーグル奨学生
カルテンポト M.
エングル T.
膜翅目における防御的微生物共生体。
Funct. Ecol. 2014; 28: 315-327
論文で見る
スコープス (118)
クロスリファレンス
グーグル奨学生
バークレイ V.C.
シム D.
チャン・B.H.K.
ネル L.A.
ラバー M.A.
ベル A.S.
アンダース R.F.
リード A.F.
齧歯類マラリア原虫チャバディに対する血液期ワクチン接種の進化的帰結。
PLoS Biol.
論文で見る
スコパス (43)
クロスリファレンス
グーグル学者
デュアール R.M.
リン X.B.
Zheng J.
マルティーノ M.E.
グルニエ T.
ペレス-ムニョスM.E.
ルリエ F.
ゲンツレ M.
ウォルター J.
変遷するライフスタイル:ラクトバチルス属の進化と自然史。
FEMS Microbiol. Rev. 2017; 41: S27-S48
論文で見る
スコープス (200)
PubMed
クロス
グーグル奨学生
マッケンジー V.J.
クーネマンJ.G.
ハリス R.N.
野生動物の疾病緩和ツールとしてのプロバイオティクス:食品生産と医療からの洞察。
Ann. N. Y. Acad. Sci. 2018; 1429: 18-30
論文で見る
スコープス (37)
PubMed
クロス
グーグル奨学生
ホイトJ.R.
ラングウィッグK.E.
ホワイトJ.P.
カーラッカ H.M.
レデル J.A.
パリーゼ K.L.
フリック W.F.
フォスター J.T.
キルパトリック A.M.
白鼻症候群からコウモリを守るためのプロバイオティクス細菌の野外試験。
Sci. Rep. 2019; 99158
論文で見る
スコープス(48)
クロスリファレンス
グーグル奨学生
ブレッツ M.C.
ルードン A.H.
ベッカー M.H.
ベル S.C.
ウッドハムズD.C.
ミンビオレK.P.C.
ハリス R.N.
バイオオーグメンテーションによる両生類ツボカビ症の軽減:効果的なプロバイオティクスの特徴と、その選択と使 用の戦略。
Ecol. 2013; 16: 807-820
論文で見る
日本生物工学会誌
PubMed
クロス
グーグル奨学生
クロス M.L.
微生物対微生物:プロバイオティック乳酸菌によって生成される免疫シグナルと微生物病原体に対する防御におけるその役割。
FEMS免疫。Med. Microbiol. 2002; 34: 245-253
論文で見る
パブコメ
クロスレフ
グーグル奨学生
ウーヴェハンド A.C.
フォーステンS.
ヒバードA.A.
ライラA.
シュタールB.
抗生物質耐性を防ぐためのプロバイオティクスアプローチ。
アン。Med. 2016; 48: 246-255
論文で見る
スコープス (106)
PubMed
クロス
グーグル奨学生
Stiernagle T.
線虫の維持。
WormBook. 2006; 2006: 1-11
記事で見る
Google Scholar
ベガ N.M.
ゴアJ.
Stochastic assembly produces heterogeneous communities in the Caenorhabditis elegans intestine.
PLoS Biol.
論文で見る
スコープス(77)
クロス
グーグル奨学生
コールド・スプリング・ハーバー
線虫増殖培地(NGM)。
Cold Spring Harb. Protoc.、2008
https://doi.org/10.1101/pdb.rec11474
論文で見る
クロス
グーグル・スカラー
ハ D.
クチマ S.L.
トゥール G.A.O.
緑膿菌の遊泳運動性のプレートベースアッセイ。
Methods Mol. Biol.
論文で見る
スコパス (74)
PubMed
クロス
グーグル奨学生
陳 S.
Zhou Y.
Chen Y.
Gu J.
Fastp:超高速オールインワンFASTQプリプロセッサ。
Bioinformatics. 2018; 34: i884-i890
論文で見る
スコープ(8294)
PubMed
クロスリファレンス
グーグル奨学生
イルミナ
bcl2fastq:Bclファイルをベースコールに変換するためのイルミナ独自のソフトウェア。
2013
論文で見る
Google Scholar
Wick R.
Porechop:ONTテクノロジーのQCおよびアダプタートリミングのためのオープンソースソフトウェア。
2018
https://github.com/rrwick/Porechop
記事で見る
グーグル・スカラー
ウィック R.R.
ジャッド L.M.
ゴリー C.L.
ホルト K.E.
Unicycler: 短鎖および長鎖シーケンスリードからの細菌ゲノムアセンブリの解析。
PLoS Comput. Biol.
論文で見る
スコープス (3851)
クロスリファレンス
グーグル奨学生
シュベンガー O.
イェロネクL.
ディークマンM.A.
ベイバース S.
ブロム J.
ゲーズマン A.
Bakta: アラインメントフリー配列同定による細菌ゲノムの迅速かつ標準化されたアノテーション。
Microb. Genomics. 2021; 7000685
論文で見る
クロスリファレンス
グーグル奨学生
Li H.
ハンザカー B.
ワイソーカーA.
フェネル・T.
ルアンJ.
ホーマー N.
マース G.
アベカシスG.
ダービン R.
Sequence Alignment/MapフォーマットとSAMtools。
Bioinformatics. 2009; 25: 2078-2079
論文で見る
筑波大学
PubMed
クロスリファレンス
グーグル奨学生
Rコアチーム
R: 統計計算のための言語と環境.
2021
https://www.r-project.org/
記事で見る
グーグル・スカラー
Deatherage D.E.
バリック J.E.
breseqを用いた次世代シーケンスデータからの実験室内で進化させた微生物における変異の同定。
Methods Protoc. 2014; 1151: 165-188
論文で見る
Google Scholar
Kofler R.
パンデイ R.V.
Schlötterer C.
PoPoolation2:プールされたDNAサンプルのシーケンス(Pool-Seq)を用いた集団間の分化の同定。
Bioinformatics. 2011; 27: 3435-3436
論文で見る
論文掲載
パブコメ
クロスリファレンス
グーグル奨学生
シャーマン B.T.
ハオ M.
Qiu J.
Jiao X.
Baseler M.W.
レーン H.C.
Imamichi T.
Chang W.
DAVID:遺伝子リストの機能濃縮解析と機能アノテーションのためのウェブサーバー(2021年更新)。
Nucleic Acids Res. 2022; 50: W216-W221
論文で見る
遺伝子発現情報
PubMed
クロスリファレンス
グーグル奨学生
金久正明
後藤慎一郎
KEGG:Kyoto Encyclopedia of Genes and Genomes(京都遺伝子ゲノム百科事典)。
Nucleic Acids Res. 2000; 28: 27-30
論文で見る
PubMed
クロスフィルム
グーグル奨学生
Felsenstein J.
系統推論パッケージ(PHYLIP)。バージョン3.5。
(ワシントン大学、シアトル)1993
論文で見る
Google Scholar
パラディ E.
シュリープ K.
Ape 5.0: Rによる最新の系統解析と進化解析のための環境。
Bioinformatics. 2019; 35: 526-528
論文で見る
スコープス (3836)
PubMed
クロスリファレンス
グーグル奨学生
論文情報
出版履歴
発行 2024年3月1日
受理 受理:2024年2月7日
改訂版受理 2023年12月18日
受理:2023年12月18日 受理日:2023年10月9日
出版段階
インプレス、修正校正
識別
DOI: https://doi.org/10.1016/j.cub.2024.02.015
著作権
© 2024 The Author(s). エルゼビア社発行
サイエンスダイレクト
ScienceDirectでこの記事にアクセスする
図
図サムネイルfx1
グラフィカルアブストラクト
サムネイルgr1
図1宿主微生物叢は不完全な免疫防御を提供する
図のサムネイルgr2
図2微生物叢による不完全な免疫により、より病原性の強い病原体が選択される
図サムネイルgr3
図3微生物叢からの不完全な免疫により病原体の分子進化が抑制される
図サムネイルgr4
図4微生物叢が誘導する宿主防御が病原体の進化経路を変える
関連記事
広告
ライフ&メディカルサイエンスジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノム学
細胞宿主と微生物
細胞代謝
細胞レポート
セルレポーツ医学
細胞幹細胞
細胞システム
カレントバイオロジー
発生細胞
免疫
医学
分子細胞
ニューロン
構造
アメリカン・ジャーナル・オブ・ヒューマン・ジェネティクス (パートナー)
生物物理学雑誌 (パートナー)
生物物理レポート (パートナー)
Human Genetics and Genomics Advances (パートナー)
分子植物 (パートナー)
分子療法 (パートナー)
分子治療法と臨床開発 (パートナー)
分子療法核酸 (パートナー)
分子療法腫瘍学 (パートナー)
Plant Communications (パートナー)
幹細胞レポート (パートナー)
生化学の動向
癌の動向
細胞生物学の動向
生態学と進化の動向
内分泌学・代謝学の動向
遺伝学の動向
免疫学の動向
微生物学の動向
分子医学の動向
神経科学の動向
寄生虫学の動向
薬理学の動向
植物科学の動向
物理科学・工学ジャーナル
セルレポート 物理科学
化学
化学触媒
デバイス
ジュール
物質
化学の動向
学際ジャーナル
セルリポーツ・メソッド
セルリポーツ・サステイナビリティ
Heliyon
アイサイエンス
ワン・アース
パターン
STARプロトコル
ネクサス(パートナー)
イノベーション(パートナー)
バイオテクノロジーの動向
認知科学の動向
著者
論文投稿
複数ジャーナル投稿
STARメソッド
プレビュー - プレプリント
査読者
査読者向け情報
ニュース&イベント
ニュースルーム
細胞シンポジウム
コンソーシアムハブ
ウェビナー
ラボリンク
マルチメディア
セルプレスポッドキャスト
セルプレスビデオ
カラーリングとコミック
フィギュア360
セル画ショー
研究篇
セルプレスについて
セルプレスについて
オープンアクセス
COVIDハブ
持続可能性
インクルージョンと多様性
コンタクト
お問い合わせ
ヘルプ&サポート
採用情報
セルプレス採用情報
サイエンティフィックジョブボード
アクセス
登録する
請求
今すぐ読む
司書に推薦する
出版アラート
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
Nucleusコレクション
スナップショット・アーカイブ
インフォメーション
広告主の皆様へ
リクルーターの方へ
図書館員の方へ
プライバシーポリシー
ご利用条件
アクセシビリティ
本サイトのコンテンツは、あらゆる分野の医療従事者および研究者を対象としています。
当サイトでは、サービスの提供・向上およびコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を更新するには、このサイトのクッキー設定をご覧ください。
このサイトのすべてのコンテンツ: Copyright © 2024 Elsevier Inc.、そのライセンサー、および寄稿者。
テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビア社に帰属します。
すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条件が適用されます。
プライバシーポリシー 利用規約 アクセシビリティ ヘルプ&サポート お問い合わせ
RELX
この記事が気に入ったらサポートをしてみませんか?