最も処方されている薬剤が微生物叢-腸-脳軸に及ぼす潜在的影響:総説

本文へスキップ記事へスキップ
エルゼビアのロゴ
ブレイン・リサーチ・ブレティン
オンラインで入手可能 2024年1月18日, 110883
In Press, Journal Pre-proofこれは何ですか?
レビュー
最も処方されている薬剤が微生物叢-腸-脳軸に及ぼす潜在的影響:総説
https://www.sciencedirect.com/science/article/pii/S0361923024000169
著者リンク オーバーレイパネルを開くKirti Garg, M. Hasan Mohajeri
もっと見る
概要
共有
引用
https://doi.org/10.1016/j.brainresbull.2024.110883
権利とコンテンツの取得
クリエイティブ・コモンズ・ライセンス
オープンアクセス
ハイライト
日常的に処方される薬剤(メトホルミン、スタチン、PPI、非ステロイド性抗炎症薬)は、ディスバイオシスを引き起こす可能性がある。
これらの薬剤は、互いのディスバイオシスを増強または相殺する可能性がある。
薬剤誘発性ディスバイオシスは、MGBAを通じて脳疾患に影響を及ぼす可能性がある。
その影響は、異なる細菌が産生する、あるいは修飾された代謝産物を通じて起こるかもしれない。
多剤併用高齢者が薬剤性ディスバイオシスの影響を最も受ける可能性がある。
要旨
微生物叢-腸-脳軸(Microbiota-Gut-Brain Axis:MGBA)と名付けられた、腸内常在菌とその代謝産物を介した薬剤誘発性ディスバイオシスと脳疾患への影響との関連は、まだほとんど解明されていない。本総説では、一般的に処方されている薬剤(メトホルミン、スタチン、プロトンポンプ阻害薬、非ステロイド性抗炎症薬、抗うつ薬)が腸内細菌叢に及ぼす影響について、主要な脳疾患(うつ病、多発性硬化症、パーキンソン病、アルツハイマー病)における細菌集団の変化と比較しながら調査している。本報告の目的は、薬剤がMGBAを介して脳疾患の発症と進行に影響を及ぼすかどうかを探ることである。中心的な知見は、探索されたすべての薬剤がディスバイオシスを誘導することを示している。これらのディスバイオシスのパターンは脳疾患と関連していた。脳疾患への影響は細菌分類群によって異なり、おそらく直接的な影響または細菌の代謝産物を介したものであろう。各薬剤は細菌の存在量に正の変化と負の変化の両方を誘導し、相殺効果を示した。さらに、上記の薬剤は類似した効果を示したことから、併存患者が一緒に服用した場合、脳疾患に対する互いの効果を相殺または増強する可能性が示唆された。結論として、細菌種とその存在量の相互作用は、個々の薬剤や細菌株よりも脳疾患に大きな影響を及ぼす可能性がある。薬物誘発性ディスバイオシスと脳疾患発症への影響をより深く理解し、脳関連疾患患者に対してより効果的な治療法を開発できる可能性を秘めた今後の研究が必要である。
略語
ADAlzheimer's diseaseBact2Bacteroides enterotype 2BBBblood brain barrierBMIBody max indexCDIClostridium difficile infectionCNScentral nervous systemEECsenteroendocrine cellENSenteric nervous systemGABAγ- アミノ酪酸GBA腸-脳軸GGF無胚葉GIT消化管GLP-1グルカゴン様ペプチド-1GM腸内細菌叢HPAA視床下部-下垂体-副腎軸IBD炎症性腸疾患LDL低 低密度リポタンパク質LPSリポ多糖METメトホルミンMGBA微生物叢-腸-脳軸MS多発性硬化症NSAID非ステロイド性抗炎症薬PDPパーキンソン病PPIプロトンポンプ阻害薬PYペプチドYYRCTランダム化比較試験SCFA短鎖脂肪酸SIBO小腸腸管過成長SSRI選択的セロトニン再取り込み阻害薬TLR4トール様受容体4
キーワード
薬剤誘発性ディスバイオシス細菌代謝産物メトホルミンPPINSAIDs抗うつ薬スタチンうつ病多発性硬化症パーキンソン病アルツハイマー病
細菌名の同義語
このレビューで言及された名前 NCBI ((NCBI), 2023)による同義語
ファーミキューテス属
グリチルリチン酸クロストリジウム Mediterraneibacter glycyrrhizinilyticum
ラクトバチルス・ブレビス Levilactobacillus brevis
バクテロイデーテス バクテロイデーテス門
ローソニバクター・フォセンシス ローソニバクター・アサッカロリティカス
クロストリジウム・ココイデス Blautia coccoides
ルミノコッカス・ニャバス メディタラネイバクター・ニャバス
アネロブチリカム
Eubactrium hallii Anaerobutyricum hallii
はじめに
ヒトは様々な体の部位で様々な微生物組成を宿主としているが、最も大きなものは消化管である。微生物群は代謝産物を産生し、人体の様々なプロセスに役立っている。ディスバイオーシスは、微生物叢のアンバランス、機能的組成や代謝活性の変化、あるいは局所的分布の変化をもたらすマイクロバイオームの崩壊を特徴とする。ディスバイオーシスは、肝疾患、心血管疾患、神経疾患から、がんや炎症性腸疾患(IBD)などの自己免疫疾患まで、さまざまな疾患に影響を及ぼす可能性がある。最近の研究では、腸内常在菌と脳との間の複雑な双方向コミュニケーション、特に神経疾患におけるその意味合いが強調されている(Agnihotri and Mohajeri, 2022, Knuesel and Mohajeri, 2021)。さらに、動物モデルを用いた前臨床研究では、腸内細菌組成の変化に関する知見を再現し、微生物叢が脳の病態に及ぼす影響を示すことができた。この証拠から、ヒトの腸内細菌が脳の生理・病理に寄与している可能性が示唆される(Moraisら、2021年)。生きた細菌株を用いた新しい医薬品、すなわちプロバイオティクスや、よく考えられた食事は、腸内細菌叢(GM)の完全性を回復し、これらの影響をある程度改善することが示されている(Beamら、2021年)。
幅広い健康問題を治療するために、これまでに15'000種類近くの医薬品が開発されている(Online, 2022)。2021年には、アメリカの人口の66%が処方薬を服用していた。カナダは約65%(2019年)、イギリス国民の26%以上(2020年)、オーストラリア国民の35%以上(2018年)が処方薬使用者であった(SingleCare.com、2022年)。他のOECD(経済協力開発機構)加盟国でも、一般的に使用される医薬品の多くが増加している。2000年から2019年の間に、降圧剤の消費量は65%増加し、抗糖尿病薬と抗うつ薬は2倍に、脂質改善薬はほぼ4倍に増加した(OECD iLibrary、2021年)。インドにおける市販薬のセルフメディケーションは約52%で、潜在的な好ましくない副作用のために健康リスクが高まっていることを示している(Bindu Shajan Perappadan, 2015)。
これらの薬剤の中には、糖尿病、高血圧、がん、関節炎などの慢性疾患のために処方されるものもある。世界中で最もよく使用されている薬剤は、抗糖尿病薬、心臓血管薬、鎮痛薬、プロトンポンプ阻害薬、抗うつ薬である(ClinCalc.com、2020年)。WHO(世界保健機関)は、前述のクラスに属する医薬品のうち、基本的な医療制度に不可欠なものを取り上げている(Organisation, 2021)。これらの "必須 "薬剤の多くは、(多疾患合併症患者などが)一緒に服用すると、互いの効果や副作用を打ち消し合う可能性がある。したがって、患者を最適に治療するためには、その効果と副作用の両方を知ることが非常に重要であることは直感的に理解できる。
抗生物質は人間の健康にとって重要である。しかし、抗生物質の主な副作用のひとつは、腸内細菌組成の深刻な変化である。さらに多くの研究が、抗生物質が誘発する腸内細菌異常と腸脳軸(GBA)との間に強い関係があることを示している。抗うつ薬、スタチン、非ステロイド性抗炎症薬など、他の医薬品も同様の腸内微生物変化作用を有することが報告されている(Essmat et al.) したがって、我々は「新たに発見された」抗菌特性を持つ薬剤と、微生物叢-腸-脳軸(MGBA)に影響を与えるそのメカニズムとの間に関連があると仮定している。
この総説は、糖尿病や心血管疾患などの他の病的疾患のために毎日服用される薬物によって引き起こされる薬剤性腸内細菌叢異常症とその推定されるメカニズムに関する知識を得ることを目的としている。そして、薬物誘発性ディスバイオーシスを外挿することで、MGBAに影響を及ぼす可能性のある影響について将来の展望を探る。そこで本稿では、メトホルミン(MET)、プロトンポンプ阻害薬(PPI)、スタチン、非ステロイド性抗炎症薬(NSAIDs)、抗うつ薬によって生じる腸内細菌叢(GM)の変化についてまとめる。順次、成人において診断される主要な神経疾患および神経変性疾患であるうつ病、多発性硬化症(MS)、パーキンソン病(PD)、アルツハイマー病(AD)について、微生物叢-腸-脳軸に影響を及ぼす作用機序を仮定する。図S1、S2、S3、S4、S5、S6、S7、S8、S9、S10の補足資料の項に、言及した細菌名が分類学的順序でまとめられている。
材料と方法
PubMedは2つの異なる観点から論文を検索した。まず、メトホルミン、プロトンポンプ阻害薬、非ステロイド性抗炎症薬、スタチン、SSRI、抗うつ薬と腸内細菌叢に関する論文を検索した。除外キーワードは、抗生物質、プロバイオティック、プレバイオティックであった。検索結果は2022年8月までであった(図1)。定性的・定量的解析のため、本レビューでは動物実験よりもヒトの研究に重点を置いている。
図1
ダウンロード 高解像度画像ダウンロード(262KB)
ダウンロード フルサイズ画像のダウンロード
図1. PRISMA基準(Statement, 2020)を用いたレビュー(薬剤の検索)の系統的アプローチのフロー図(2段組みのはめ込み画像/図である)。
その後、2回目の検索では、腸内細菌叢と腸脳軸をキーワードに、それぞれの疾患と関連する論文を検索した: うつ病、アルツハイマー病、パーキンソン病、多発性硬化症である。ここでは、検索範囲を過去6年間(2017~2023年)、「ヒト」に限定した。PubMedの詳細検索で除外したキーワードは、プロバイオティクス、抗生物質、出生前、母体、過敏性腸症候群、食事、アルコール、乳児、小児、思春期、がん、化学療法、肝硬変、肥満であった(図2)。これらのキーワードは、スクリーニングの際に上記の薬剤に関する論文を除外するためにも使用された。
図2
ダウンロード 高解像度画像ダウンロード(237KB)
ダウンロード フルサイズ画像のダウンロード
図2. PRISMA基準(Statement, 2020)によるレビューの系統的アプローチ(疾患の検索)のフロー図(これは2列のフィッティング画像/図である)。
冗長な論文、英語で発表されていない論文、レビューされた薬剤で治療されていない疾患に関する研究コホートがある論文、信頼できる対照セットがない論文、動物ベースの論文しかない論文、機関論文を通じたアクセスが認められていない論文は考慮されなかった。選択された論文の参考文献に記載された報告は、必要に応じて追加された。
本論文の図版は、それぞれの凡例に記載されたライセンス番号のもと、BioRenderソフトウェアで作成した。
腸内細菌叢
人体の様々な部位には様々な微生物が生息しており、各部位に固有の細菌叢を形成している。一般的にマイクロバイオームは、細菌、ウイルス、古細菌、寄生虫、真菌、真核生物から構成されている(Adak and Khan, 2019, Beam et al., 2021, Liang et al., 2018, Sidhu and van der Poorten, 2017)。腸内細菌は2億3,200万個以上の遺伝子をコードしており、ヒトの代謝に不可欠な役割を担っていることが示唆されている(Eicher and Mohajeri, 2022, Morais et al.) 腸内には、これらの微生物のうち最も多くの微生物が生息しており、その数は100兆個、約5000種、重さは約2kgにもなる(Beam et al., 2021, Gomaa, 2020, Sidhu and van der Poorten, 2017)。本稿では、消化管、すなわち胃と腸の細菌組成に焦点を当てる。
腸内細菌叢(GM)は母親の胎内や出生時に獲得された後、年齢やストレスなどのさまざまな生活経験によって、その個体間組成は生涯にわたって変化する(Adak & Khan, 2019)。腸内細菌叢(GM)は、加齢、地理的位置、民族性、食事、薬物、毒素、病気などの環境やライフスタイルの要因によって影響を受ける(Adak and Khan, 2019, Cryan et al., 2019, Koo et al.) 双子ペアの糞便サンプルに基づく英国での研究も、宿主の遺伝がGMに影響を及ぼす役割を指摘している(Adak & Khan, 2019)。
神経変性疾患、心血管疾患、肝疾患、自己免疫疾患などの健康や疾患において、遺伝子組み換えが重要な役割を果たす可能性を論じる証拠が増えている。GMはまた、2型糖尿病、高血圧、メタボリックシンドロームなどの生活習慣病、あるいは神経疾患やがんなどの生命を脅かす疾患に対して、薬剤の生物学的利用能とその有効性において明らかな役割を果たしている可能性がある(Weiss & Hennet, 2017)。これらの発見は、人体とマイクロバイオームが共生関係にあるという考えに火をつけた。後者は、直接的または間接的なメカニズム(Morais et al. これらのメカニズムには、栄養素の代謝、強力な宿主免疫系と腸管バリアの完全性の創出、酵素、ビタミン(ビタミンKなど)、神経伝達物質(セロトニンなど)の産生、有益菌種と日和見菌種の間の恒常性の維持、すなわちユビオシスが含まれる(Beam et al.)
ユビオシスは微生物組成における恒常性状態を定義する。ディスバイオーシス」という用語(1985年にBaiによって初めて使用された;Otani et al. ディスバイオーシスは全身性の炎症を引き起こし、メタボリックシンドローム、2型糖尿病、神経疾患、自己免疫疾患などの疾患の発症につながる。ディスバイオーシスは、環境要因や生活習慣要因によって誘発されることがある(Beamら、2021年)。
3.1. 腸内細菌叢の構成と検査
消化管の様々な微小環境により、消化管の様々な部位で異なる細菌集団が増殖する。ファーミキューテス属、バクテロイデーテス属、アクチノバクテリア属、フソバクテリア属、プロテオバクテリア属、ベルコミクテリア属、シアノバクテリア属が7つの門を形成している。成人では、腸の90%がバクテロイデーテス(主にグラム陰性でプレボテラ)、ファーミキューテス(一般にグラム陽性でクロストリジウム、ルミノコッカス、ユーバクテリウムが主体)で構成されている(Adak and Khan, 2019, Gomaa, 2020, Liang et al., 2018, Pellegrini et al.) 特に、バクテロイデーテス(Bacteroidetes)とファーミキューテス(Firmicutes)の比率は、炎症性マイクロバイオーム(pro-inflammatory microbiome)へのシフトを伴う炎症プロセスにおいて重要な役割を果たしている(Adak and Khan, 2019, Gomaa, 2020, Liang et al., 2018, Pellegrini et al., 2018, Wang et al., 2021, Weiss and Hennet, 2017)。胃と腸に存在する腸内細菌組成の概要を(図3)に説明する。これらの細菌コロニーをその特徴や生息域に基づいて区別するために、糞便サンプル、生検、吸引が検査される。尿サンプルと血液サンプルは、その代謝産物を介して細菌コロニーを区別するためにチェックされる。16 s rRNAゲノム配列決定法とショットガン配列決定法により、培養に依存する属と培養に依存しない属と種をそれぞれ同定することができる(Adak & Khan, 2019)。
図3
ダウンロード: 高解像度画像のダウンロード(313KB)
ダウンロード フルサイズ画像のダウンロード
図3. 胃と腸の消化管における腸内細菌叢組成の概要。一般に、上部消化管から下部消化管にかけて、好気性菌から嫌気性菌への移行が見られる。消化管(GIT)の最初の部分である胃には、主に5大菌門であるファーミキューテス(Firmicutes)、バクテロイデス(Bacteroides)、アクチノバクテリア(Actinobacteria)、フソバクテリア(Fusobacteria)、プロテオバクテリア(Proteobacteria)と、ストレプトコッカス(Streptococcus)、プレボテラ(Prevotella)、ベイロネラ(Veillonella)、ロチア(Rothia)、ヘモフィルス(Haemophilus)属の耐酸性細菌種が生息している。次に小腸は、十二指腸、空腸、回腸の3つの部分からなる。十二指腸から回腸に向かうにつれて、細菌種とその数は変化する。十二指腸のファーミキューテスと放線菌から、空腸のグラム陽性好気性菌と通性嫌気性菌へと移行する。回腸は好気性菌と嫌気性菌、グラム陰性菌からなる。盲腸と結腸は主にファーミキューテスとバクテロイデーテスからなる。(イラストはBioRender.com(Software, 2023)で作成: OR25UMC72C)(これは2列のフィッティング画像/図である)。
微生物叢-腸-脳軸
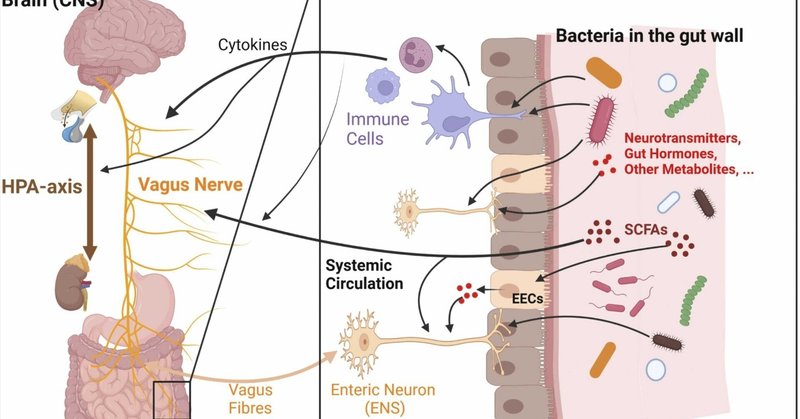
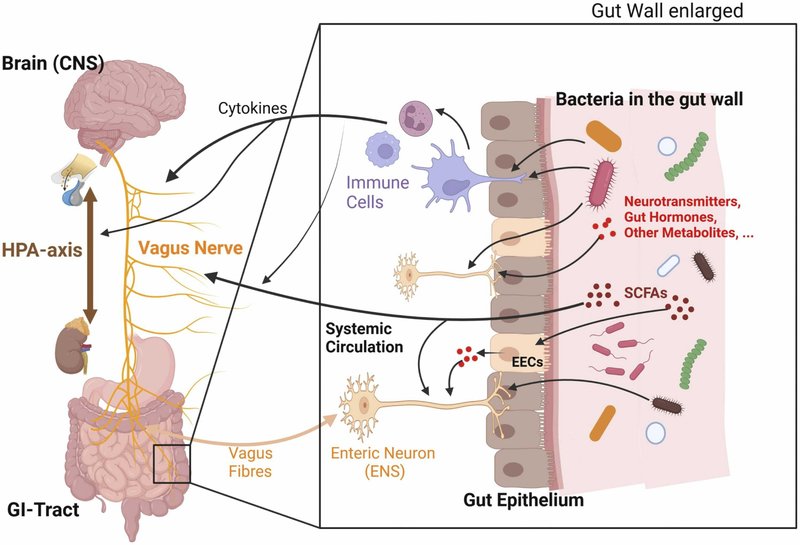
腸-脳軸は、腸神経系(ENS)と中枢神経系(CNS)のつながりを定義する。研究が進むにつれ、微生物叢がこの軸において重要な役割を果たしていることが明らかになり、腸-脳軸は微生物叢-腸-脳軸(MGBA)へと拡張された。MGBAは、腸と脳の生理学的側面に影響を与える、腸と中枢神経系間の双方向コミュニケーション経路である。以来、このコミュニケーションにはさまざまな経路が関与していることが報告されている(図4)。GMは、神経伝達物質や代謝産物の細菌産生(いわゆる化学的シグナル伝達)、迷走神経、免疫系、あるいは視床下部-下垂体-副腎軸(HPAA)を含む神経内分泌系を介して脳に影響を及ぼす可能性がある(Chakrabartiら、2022年、Cussottoら、2018年)。
図4
ダウンロード: 高解像度画像のダウンロード(352KB)
ダウンロード フルサイズ画像のダウンロード
図4. 微生物叢-腸-脳軸(MBGA)の双方向コミュニケーションに関する概略図。腸内には多くの細菌株が生息しており、これらの細菌株は脳と直接的または間接的にコミュニケーションをとることができる。矢印は、関係する異なるステーション間の個々の経路のシグナル伝達方向を示す。神経伝達物質、サイトカイン、腸内ホルモンなど、さまざまなニューロン、免疫、内分泌メディエーター(指定されたメディエーターの順に記載)が、MGBAの双方向経路の伝達者である。放出されたサイトカインは、免疫細胞を通じて細菌を認識した後、直接脳に影響を与える。サイトカインと迷走神経は、視床下部-下垂体-副腎軸(HPA軸)にも影響を与える。細菌はまた、腸管ニューロン上のtoll様受容体-4(TLR4)を誘発するリポ多糖(LPS)を介して、腸管ニューロンと直接コミュニケーションすることもできる。間接的な経路としては、神経伝達物質や短鎖脂肪酸(SCFA)など、腸内細菌が産生する代謝産物がある。これらの代謝産物は全身の血液循環に入り、脳に影響を与える。さらに、代謝産物は腸の腸内分泌細胞(EEC)を刺激し、腸内分泌細胞は血液中に腸ホルモンを放出する。細菌の代謝産物や腸内ホルモンは、迷走神経に属する腸管ニューロンを介して求心性シグナルを送ることもできる。一方、脳は迷走神経線維や他の腸神経系(ENS)細胞を介して、求心性のシグナルを伝達する。迷走神経は、粘液分泌や腸の運動といった重要な側面において、腸の生理機能に影響を与えている。(イラストはBioRender.com(Software, 2023)で作成: DM25VBM0KY)(これは2段組のフィッティング画像/図である)。
GMは、化学的シグナルを通じて直接または間接的に脳と相互作用する(Chakrabartiら、2022)。化学的シグナルには主に、短鎖脂肪酸(SCFA)、アミノ酸、神経伝達物質、腸内ホルモン、神経栄養因子が含まれる。化学シグナル伝達は、すべての経路を相互に接続している。これらの化学物質の多くは微生物産物であり、ニューロン経路を介して脳に影響を与える可能性がある(Morais et al.)
酢酸、プロピオン酸、酪酸は、抗炎症反応を発揮する腸内で最も豊富なSCFAである(Dalileら、2019)。SCFAは、宿主の難消化性食物繊維の発酵によって細菌が産生する飽和脂肪酸である。SCFAはある程度血液脳関門(BBB)を通過することができ(Dalile et al. SCFAの化学的シグナル伝達は、グルカゴン様ペプチド-1(GLP1)、ペプチドYY(PYY)、レプチン、グレリン、インスリンなどの特定の神経伝達物質や腸内ホルモンの分泌も調節することができる(Chakrabartiら、2022、Dalileら、2019)。
腸内細菌はSCFAとともに、アミノ酸の一種であるトリプトファンなど、様々な神経活性分子を産生または代謝している(Chakrabarti et al.) トリプトファンはBBBを通過することができ(Chakrabarti et al. したがって、BBB透過性のSCFAとトリプトファンは、脳回路に影響を与え、気分や認知を含む脳機能に影響を及ぼすセロトニンの脳内生合成と利用可能性に影響を及ぼす可能性がある(Chakrabartiら、2022、Liangら、2018)。セロトニンの90%以上は、SCFAによって活性化された腸内分泌細胞(EEC)を通じて腸管系で合成される(Chakrabartiら、2022、Eicher and Mohajeri、2022)。胃腸セロトニンは、連鎖球菌属、カンジダ属、腸球菌属、大腸菌属によっても産生される(Eicher & Mohajeri, 2022)。さらに、ドーパミンの調節は、中毒、統合失調症、パーキンソン病とも関連している(Chakrabartiら、2022年)。バチルス属やビフィドバクテリウム属など、特定の細菌がドーパミンの産生や調節に関与している可能性が研究で示唆されているため(Eicher and Mohajeri, 2022, Strandwitz, 2018)、ドーパミン作動性神経伝達の混乱は、ディスバイオシスの結果である可能性がある(González-Arancibia et al.) 同様の報告により、GMの多様性と豊かさの摂動が、セロトニン作動性、ドーパミン作動性だけでなく、GABA作動性、ノルアドレナリン作動性、グルタミン酸作動性神経伝達も形成することが示されている(Chakrabartiら、2022、González-Arancibiaら、2019)。
前述の拡散性代謝産物、腸内ホルモン、神経伝達物質は、副交感神経迷走神経反応、すなわち迷走神経(VN)も刺激する。例えば、注射した酪酸ナトリウムの拡散は、雄ラットにおいて迷走神経求心性活動電位を誘発し、横隔膜下迷走神経切開術を行うと、この活動は停止した(Dalile et al.) さらに、迷走神経求心性線維はtoll-like receptor 4(TLR4)を備えており、リポ多糖(LPS)を感知して脳を活性化することができる(Bonaz et al.) これらの知見は、腸内細菌叢と迷走神経刺激との関連を示唆している。
グラム陰性菌の菌壁に見られる免疫トリガーLPSは、腸管細胞によっても検出される。その結果、局所および全身の免疫応答カスケードが活性化される(Eicher and Mohajeri, 2022, Weiss and Hennet, 2017)。全身性免疫の調節障害は全身性炎症を促進し、さまざまな免疫カスケードやBBBなどのバリアを破壊する。その結果、脳における毒素からの保護が低下し、様々な神経障害の発症が促進される(Cryan et al.) BBBの完全性が損なわれることは、多くの神経病理学的障害に共通する特徴である(Eicher and Mohajeri, 2022, Morais et al., 2021)。マウスでは、GMの変化によりBBBの透過性が亢進している証拠がある(Moraisら、2021)。GFマウスはBBBの透過性亢進を示したが、これは酪酸塩処理によって病原体を持たないマウスのレベルまで直接逆転した(Dalileら、2019年)。さらに、透過性BBBはミクログリアの機能不全を介して神経炎症を調節する可能性がある。抗生物質を投与したGFマウスはミクログリア機能が著しく低下していたことから、微生物叢は中枢神経系におけるミクログリアの構造と機能的完全性に影響を及ぼすことが判明した(Morais et al.) ミクログリアの発現パターンの変化は、うつ病、自閉症スペクトラム障害、強迫性障害など多くの精神疾患で報告されている(Eicher & Mohajeri, 2022)。さらに、微生物叢が減少した動物や微生物叢がない動物では、ミクログリアの炎症表現型と循環SCFAs量の乱れとの間に正の相関が見られたことから、SCFAsとミクログリアの機能は相互に関連している可能性があり、MGBAを介したSCFAsのさらに別の間接的な効果を提唱している(Cryan et al.) ミクログリア細胞以外にも、アストロサイトは神経炎症を介して活性化される。慢性的な炎症やストレスは、アストロサイトの活性化を維持し、したがってグルタミン酸作動性伝達も維持することができる。活性化したアストロサイトはグルタミン酸の内在化を減少させ、グルタミン酸レベルを増加させ、結果としてグルタミン酸作動性興奮伝達を増加させることが報告されている(Leeら、2022年、Mahmoudら、2019年、Troubatら、2021年)。興奮毒性グルタミン酸伝達の亢進は、神経毒性、特にうつ病(Troubatら、2021年)、アルツハイマー病、パーキンソン病、ハンチントン病、てんかん(Leeら、2022年)などの脳障害において重要であるとみなされてきた。このことは、GMの多様性と豊かさの摂動が様々な種類の神経伝達を形成し(Chakrabarti et al., 2022, González-Arancibia et al., 2019)、GMの変化が免疫系に影響を及ぼし、それによってBBBを変化させるという、前述の同様の知見につながる。
ストレス反応の調節異常は多くの精神神経疾患に見られ、うつ病や自閉スペクトラム症はその2つの例に過ぎない。人体におけるストレスと神経内分泌経路の大きな伝達者であり調節者であるのが、視床下部-下垂体-副腎軸(HPAA)である。ストレスに対する反応としてHPAAの反応亢進を示すGFマウスにおける証拠は、GMを介したHPAAの調節を示唆し、ひいてはストレス調節におけるGMの役割を示唆している。病気行動に関与する迷走神経のシグナル伝達回路も、LPSとTLR4を介してHPAAを活性化することができる(Agirmanら、2021年)。Beldaらは、ドーパミンもまた、グルココルチコイドが関与する第一のストレス反応のためにHPAAを刺激すると論じている(Belda & Armario, 2009)。従って、ディスバイオシスと異なるMGBA経路は、視床下部-下垂体-副腎軸を変化させる。
総じて、GMはMGBAと様々な形で相互作用し、その結果、MGBAを変化させたり、影響を与えたりする上で重要な役割を果たすことが報告されている。
結果
本総説で示されたデータは、最も処方されている薬剤のいくつかであるメトホルミン、スタチン、プロトンポンプ阻害薬、非ステロイド性抗炎症薬、抗うつ薬が、その摂取により生体内異常を示すことを示している。この章では、それぞれの薬剤について、その摂取が患者のマイクロバイオームの構成に及ぼす影響を説明し、細菌量に関する最も顕著な変化を概説する。そして第6章では、薬物誘発性ディスバイオーシスと、脳関連疾患(うつ病、多発性硬化症、パーキンソン病、アルツハイマー病)で観察される脳疾患関連ディスバイオーシスとの類似性について論じ、取り上げた薬物と疾患との潜在的な相関性を仮定する。
5.1. メトホルミン
抗糖尿病薬メトホルミン(MET)は、2型糖尿病(T2DM)の第一選択薬であり、すでに60年以上にわたってT2DM患者に処方されている(C. B. Leeら、2021年)。米国だけでも、2020年に9,259万件のMET処方が報告されており(Statista.com、2022年)、糖尿病予備軍、糖尿病患者、またはインスリン抵抗性の症状に対するものである。経口投与後、小腸で吸収された後のMETの生物学的利用率は約40~60%である(via medici, 2023)。ヒトの腸内では、血漿中よりも30~300倍高い濃度で検出される(Y. Leeら、2021年)。METは、インスリンの効き目を改善し、肝糖新生を抑制し、肝臓でのグルカゴンシグナル伝達を抑制し、骨格筋でのグルコース取り込みを増加させ、体重を減少させることにより、血糖値を調節することでその効果を発揮する。新しい研究では、さまざまなタイプの腫瘍の発症やさらなる増殖を抑制することで、METの抗がん作用が示されている(Kanetoら、2021年)。さらに、METはGMとの相互作用を通じてグルコースレベルを低下させることが示されている(Zhang & Hu, 2020)。
MET、エンパグリフロジン、GLP-1受容体作動薬のような抗糖尿病薬の腸内環境を変化させる作用や、その逆の作用が、マウスモデルやヒトにおいて証明されつつある(Dengら、2022;Y. Leeら、2021;Liuら、2022)。METは糖尿病患者や糖尿病前症患者(Wuら、2020年)においてGMを変化させ、健常人と比較してGMが変化していることがすでに報告されている(Liuら、2022年)。METの腸内環境を変化させる作用は、腸内細菌叢に対する直接的な作用、および/または他の代謝産物を介した間接的な作用によるものである可能性がある。Maniarらの研究は、高インスリン血症も腸内細菌叢を変化させることを示唆している(Maniarら、2017)。したがって、METを介してインスリンを低下させることは、腸内細菌叢に間接的な変化を誘導することになる。
METによるGMとその微生物代謝産物の主な変化には、SCFAs産生菌の存在量の増加とLPSの上昇(おそらくグラム陰性菌の存在量の増加による)が含まれる(Almugadamら、2020、Maniarら、2017)。METの使用は、Akkermansia muciniphila、Bifidobacterium、B. adolescentisの存在量を増加させ、FirmicutesとBacteroidetesのフィラの存在量の比率を減少させた(Bryrupら、2019、Forslundら、2015、Muellerら、2021、Vallianouら、2019、Wuら、2017)。無作為化対照試験(RCT)において、Intestiniibacter barlettiiおよびRoseburia属(R. intestinalisおよびR. faecisの特定株を含む)の減少に伴うEscherichiaおよびRuminococcusのトルクス増加が示された(Muellerら、2021年)。健常人を対象に実施された試験では、インテスティニイバクター、クロストリジウム、ロンブチア属の相対量の減少と、エシェリヒア属の増加が示された(Y. Leeら、2021;Liuら、2022)。RCTで試験された他の細菌と比較して大きな変化は、ブラウチア属とフェカリバクテリウム属の増加によって示された(Tong et al.) メガモナスおよびクレブシエラ・ニューモニエは、2型糖尿病患者と比較してMET治療患者で有意に減少し(He et al. ラクトバチルス属の増加についても言及されている(Almugadamら、2020、Tongら、2018、Wuら、2017)。これらのデータは、細菌がその代謝産物および/または機能的特性を介して、2型糖尿病の血糖値を下げるというMETの効果を達成するのを助ける可能性があることを示している(表1)。
表1. メトホルミン(MET)で治療された患者において、メトホルミン(MET)によって誘発された細菌の変化が、より高い(↑、緑色)またはより低い(↓、オレンジ色)存在量で見つかった。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意である。両方の矢印(↑↓)が描かれたグレー色のセルは、高い存在量と低い存在量の両方で有意な変化があった。より高い有意水準(例:p<0.01)を持つソースは太字で示されている。矢印のついていないセルは、有意性のない相対的な変化のみを示している。
画像1
METによるGMの変化のため、この薬剤はIBDのような自己免疫疾患の治療薬としても研究されており、および/または他の腸内環境を変える薬剤の効果を元に戻すために使用されている(Wang, Huang, et al., 2021)。Wangらは、主にインスリン抵抗性や高血糖などの様々な経路を介して体重増加を引き起こし、神経活性物質(神経伝達物質や神経ペプチドなど)に影響を及ぼす抗精神病薬誘発性の代謝機能障害をMETが改善する可能性について論じている(Wang, Huang, et al.) 抗精神病薬で治療したマウスは、マイクロバイオームが枯渇したマウスよりも体重増加が大きかったことから、体重管理におけるGMの役割が示唆された。抗精神病薬を投与したマウスでは、乳酸桿菌やアッケマンシアなど多くの細菌集団が減少していた(Wang, Huang, et al.) 一方、大腸菌やビフィズス菌は増加していた。特にMETは、A. muciniphilaでも示されているように、体重増加を減少させることが示されている。後者は、全身性炎症を緩和するプロバイオティクス治療法として提案されている。したがって、METは腸内細菌叢を介して抗精神病薬使用の副作用を改善する可能性がある(Wang, Huang, et al., 2021)。
ヒトを対象とした研究はコホート規模が小さく(被験者5~25人)、まだ十分な量が存在しないため、未発表のデータは曖昧である。しかし、マウスモデルでの研究では、METとGMの相互作用が豊富に確認されている。Wuらは、ヒト患者のMET処理糞便移植を受けたマウスモデルで、ヒトの報告と同じ細菌変化を示し、METによるGMの変化を示唆した(Wuら、2017)。
5.2. スタチン
スタチンは、HGM CoA還元酵素コレステロール産生酵素を阻害することにより、低比重リポタンパク質(LDL)を低下させるために広く使用されている薬物クラスである。スタチンは心臓病や動脈硬化を予防するために最も使用されている心血管系治療薬の一つである。2億人以上の人々が様々な種類のスタチンを単独または組み合わせて服用している(Medicine, 2022)。腸内細菌叢に対するスタチンの影響を明らかにするためにヒトで行われた研究はそれほど多くなく、そのデータには一貫性がない。例えば、ロスバスタチンはパワー不足のヒト試験では有意な変化を示さなかった。一方、ロスバスタチンは腸内細菌叢の機能的可能性、すなわち細菌代謝産物のレベルにおいて有意な変化を引き起こした(Kummen et al.、2020)。あるメタアナリシス研究では、スタチンは腸内細菌型Bact2(バクテロイデス2)を減少させることにより抗炎症作用を示すことが示された(Libby, 2020, Lim, 2020, Vieira-Silva et al. スタチン治療を受けた患者では、Bact2のレベルが低く、Bacteroidesのレベルが低いことが観察された。この結果をさらに解釈すると、スタチン治療下ではファエカリバクテリウムも増加するか、バクテロイデスとファエカリバクテリウムの比率が低下し、Bact2が低下する可能性がある。スタチン治療はまた、酪酸産生マイクロバイオームの増加とも関連しているようである(Reichel & Knauf, 2021)。具体的には、アトルバスタチン治療を受けた高コレステロール血症患者では、未治療の高コレステロール血症患者と比較して、Ruminococcaceae、Verrucomicrobiaceae、A. muciniphila、Ruminococcus sp.、Oscillospira spp.、Faecalibacterium spp.の存在量が増加していた。バクテロイデス属、Faecalibacterium prausnitzii属、Akkermansia muciniphila属のような炎症促進種は減少した。Oscillospira属(抗炎症性細菌)、ファーミキューテス属、プロテオバクテリア属、Desulfovibrio属、Klebsiella属、Streptococcus属、Collinsella属などの日和見菌属も、未治療の高コレステロール血症患者と比較して、減少した常在菌で測定された。さらに、Bilophila wadsworthiaとBifidobacterium bifidum(胆汁酸関連種)の相対的減少も同じ比較で観察された。B. doreiやB. uniformisなどのバクテロイデスの異なる種における存在量の増加、同時にB. vulgatusやB. ovatusの増殖低下も、スタチン投与後に観察された(Khan et al.) これらの結果は、スタチンによるGMの変化を示しており、さらにGMを介してGBAに影響を与える可能性がある(表2)。
表2. スタチン-誘導された細菌の変化は、患者においてより高い(↑、緑色で着色)またはより低い(↓、オレンジ色で着色)存在量で認められた。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意である。矢印のついた色のついていないセルは、有意差のない相対的な変化のみを表す。
画像2
5.3. プロトンポンプ阻害薬
プロトンポンプ阻害薬(PPI)は、消化管における胃の状態を予防するために使用される。ほとんどの人、特に高齢者は、鎮痛剤や抗うつ剤などの他の薬と一緒にPPIを処方され、胃への副作用を打ち消している。PPIの作用は主に胃をターゲットにしているにもかかわらず、多くの研究が腸内細菌叢の変化を示唆している。PPIは、ヒトにおいて最も研究されている薬剤の一つである。マイクロバイオームによって誘発されるヒトの代謝への影響には、口腔、胃、小腸における細菌の過剰増殖(SIBO)、腸内クロストリジウム・ディフィシルの増加(Clooney et al、 2015, Koo et al., 2019)やサルモネラ感染症、肝性脳症、自然細菌性腹膜炎、市中肺炎、炎症性腸疾患における有害な結果、および機能的経路の変化(Naito et al., 2018)。これらの疾患や変化の多くは、PPIの使用による二次的なディスバイオシスの結果であると言われている。一般に、バクテロイデーテス(Bacteroidetes)属の菌種が少なく、ファーミキューテス(Firmicutes)属の菌種が多いことが報告されている(Clooney et al.、2016、Freedberg et al.、2015、Jackson et al.、2016)。
GITにおける特異的なdysbiosisは以下の通りである: Bacilliクラス、Lactobacillales目、Streptococcaceae科、Streptococcus属およびVeillonella属、Streptococcus vestibularisおよびVeillonella disparの存在量が有意に増加する(Kooら、2019)。腸内細菌科はPPI使用と負の相関があった(Koo et al.) Naito et al. は、バクテロイデーテスBacteroidetesバクテロイデス科、オドリバクター科、連鎖球菌科、ルミノコッカス科(Lachnospiraceae)、Megasphaera、 ActinomycesとGranulicatellaが減少し、Faecalibacterium、SMB53、Clostridium、Turicibacter、Slackia、Defluviitalea、未分類のDehalobacteriaceae、Oribacteriumが減少した(Naito et al. , 2018). 別の研究では、機能性ディスペプシア発症者では粘液に関連するナイセリア、ポルフィロモナス、セレノモナス、ヘモフィルス、フソバクテリウムの存在量が対照群と比較して低く、PPI使用後は対照群と機能性ディスペプシアでプレボテラが減少すると結論している(Wauters et al.) ナイセリアはPPI中止後も増加しており、薬剤使用後も微生物の変化は持続することが示唆された(Wauters et al.) さらに、HoldemaniaやBlautia、Granulicatella、Rothia、Doreaなどの他の細菌群やClostridiumクラスターXIVaおよびXIVbがPPI使用者で濃縮されていた(Clooney et al.) 種レベルでは、Holdemania filiformisが増加した。Clostridium glycyrrhizinilyticum(Clostridium cluster XIVa内)、Rothia mucilaginosaは有意に増加し、Pseudoflavonifractor capillosus(Clostridiales科)は減少した(Clooney et al.) Streptococcus parasanguinisとStreptococcus salivariusも同様に増加したが、有意なレベルではなかった(Clooney et al.) さらに、クロストリジウム・ディフィシル(Clostridium difficile)感染症は、連鎖球菌科(Streptococcaceae)と腸球菌科(Enterococcaceae)の集団の変化と関連している(Freedbergら、2015年)。
PPIは胃や逆流の問題を抱えている人に投与されるため、ある研究では逆流性食道炎患者のPPI治療前後の糞便微生物叢組成のみを調べた。そこでは、治療中の異なる時間間隔で、L. gasseriのサブグループ、L. reuteri、L. ruminisにL. fermentumとL. brevisを加えたようなLactobacillus属(通性嫌気性菌)が治療後に有意に増加していた(Hojo et al.) Streptococcus属、Enterobacteriaceae以下の通性嫌気性菌数、Staphylococcus属(同じく通性嫌気性菌)も同様である(Hojo et al.) C. jejuniやL. reuteriなどの細菌群を経口投与すると、迷走神経求心性シグナルの変化が見られ、それぞれ迷走神経切断マウスで元に戻るという研究がある(Cryanら、2019、Moraisら、2021)。
前述の細菌群の多くは口腔、咽頭、鼻腔で発見されており、PPIの使用による胃酸の減少によって細菌が腸管下部へ移動することが示唆されている(Hojo et al.) PPI使用による糞便サンプルでは、腸管遠位部と腸管上部とのコロニー形成が、Bacillales目(例.Staphylococcaceae)、Lactobacillales(例えば、Enterococcaceae、Lactobacillaceae、Streptococcaceae)およびActinomycetales(例えば、Actinomycetaceae、Micrococcaceae)、PasteurellaceaeおよびEnterobacteriaceae科、Veillonella属の複数の分類群の増加によって示された。減少した分類群には、ビフィズス菌科、ルミノコッカス科、ラクノスピラ科、軟体動物などが含まれる(Macke et al.、2020)。健康な双生児の研究でも同様のことが観察され、咽頭および口腔微生物叢が下部腸にコロニー形成されることによってディスバイオシスが誘導され(Jackson et al.
PPIの使用は、細菌集団のさらなるシフトを引き起こす。例えば、PPI治療後、遠位腸は上部腸の微生物叢にコロニー形成されることが示されている(Mackeら、2020年)。全体的に微生物多様性が低く、細菌種の個体数が減少しており、上部消化管では腸内常在菌の存在量が高いことが確認されている(Freedbergら、2015年、Jacksonら、2016年)。小腸細菌過剰増殖(SIBO)は後者の一例であり、Streptococcus属、Escherichia属、Klebsiella属、Bacteroides属、Lactobacillus属、Enterococcus属、Veillonella属が増加し、Bifidobacteria属とActinobactericaceaeが減少する(Fujimori, 2015)。腸の副交感神経刺激の減少は、小腸内細菌過剰増殖(SIBO)のような細菌の転位と関連している。細菌集団の変化を表3にまとめた。
表3. プロトンポンプインヒビター(PPI)-誘導された細菌の変化は、PPIで治療された患者において、より高い(↑、緑色)またはより低い(↓、オレンジ色)存在量で認められた。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意であった。両方の矢印(↑↓)が描かれたグレー色のセルは、高い存在量と低い存在量の両方で有意な変化があった。より高い有意水準(例:p<0.01)を持つソースは太字で示されている。矢印のないセルは、有意性のない相対的な変化のみを示している。
画像3
さらに、機能性ディスペプシア、肝硬変、血液透析、関節リウマチ、がん患者では、レンサ球菌属の特異的増加が認められる(Bajajら、2018、Linら、2021)。PPIは、特にヘリコバクター・ピロリ菌の感染において、消化不良を鎮めるために使用される。しかし、レンサ球菌の量を増加させるため、消化不良症状が持続する可能性がある。肛門側胃炎患者から採取した生検では、レンサ球菌の過剰増殖が認められ、レンサ球菌と消化不良との関連が確認されている(Minalyan et al.)
まとめると、非ステロイド性抗炎症薬(NSAIDs)など他の薬剤による消化不良を予防するためにPPIを使用することは、酸産生を減少させることでその効果を発揮する。しかし、PPIは近位部の腸内細菌集団が遠位部に移動するように腸内環境を変化させる。同時に、いくつかの細菌分類群の存在量が増加または減少する。その結果、このような腸内細菌異常症は、C. difficileやサルモネラ菌のような日和見菌や有害菌による腸内感染症を引き起こす。消化不良を治療するためにPPIを摂取すると、腸内細菌叢に影響を及ぼし、細菌の存在量と菌の移動が変化する。主な影響はレンサ球菌の存在量とクロストリジウム・ディフィシル感染症(CDI)の発症率に検出された。
5.4. 非ステロイド性抗炎症薬
前述の他の医薬品群と同様、非ステロイド性抗炎症薬(NSAIDs)は、疼痛や抗炎症のために日常的に処方され、また、低用量アスピリンは心血管の健康のために血小板凝集抑制剤として処方される。NSAIDsはまた、SIBOのようなディスバイオシスとその後の結果をもたらす可能性もある(Wang, Tang, et al., 2021)。非ステロイド性抗炎症薬は腸管障害を引き起こす。後者は、腸細胞の細胞死や免疫システムの破綻による腸管バリアの低下により、グラム陰性菌が増殖し、グラム陽性菌が減少する結果である(Wang, Tang, et al.、2021)。PPIと非ステロイド性抗炎症薬は、腸内細菌異常症に複合的な役割を果たしている。NSAIDsは胃症を引き起こし、PPIはその開始を抑えることができる(Rogers and Aronoff, 2016, Utzeri and Usai, 2017, Wang et al., 2021)。NSAIDsとPPIの併用摂取は腸症を引き起こすが、これはPPIがマイクロバイオームを上部消化管から遠位管にシフトさせるためである(上記参照)(Rogers and Aronoff, 2016, Utzeri and Usai, 2017, Wang et al.) 例えば、アスピリンを投与すると腸内細菌叢の組成が変化し、プレボテラ属、バクテロイデス属、ルミノコッカス属、バルネシエラ属が増加する(Rogers & Aronoff, 2016)。さらに、セレコキシブとイブプロフェンはアシダミンコッカス科と腸内細菌科を増加させる(Rogers & Aronoff, 2016)。さらにイブプロフェンは、非使用者やナプロキセン使用者と比較して、プロピオニバクテリウム科、シュードモナス科、プニセコッカス科、リケネラ科の菌種を増加させる(Maseda & Ricciotti, 2020)。インドメタシンは、バクテロイデーテス科、プレボテラ科の増加を誘導し、プロテオバクテリア科、アルファプロテオバクテリア科、プロテオバクテリア科、根粒菌科、シュードモナド科の減少を誘導した(Maseda & Ricciotti, 2020)。同じ研究では、女性腸内ではファーミキューテス類が少なく、男性腸内では多いことから、性差による影響が示された(Maseda & Ricciotti, 2020)。比較研究によると、様々なNSAIDsが誘発するディスバイオーシスでは、腸内細菌科、アシダミンコッカス科、プロピオニバクテリウム科、シュードモナド科、プニセコッカス科、リケネラ科などの分類群が増加した(Rogers & Aronoff, 2016)。上記の結果を結論づけると、多様な細菌分類群および細菌種において、ディスバイオシスおよびNSAID腸症につながる明らかなシフトが認められる(表4)。
表4. 非ステロイド性抗炎症薬(NSAIDs)により誘発された細菌の変化は、NSAIDsを投与された患者において、高い(↑、緑色)または低い(↓、オレンジ色)存在量で認められた。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意であった。
画像4
5.5. 抗うつ薬
抗うつ薬、例えば選択的セロトニン再取り込み阻害薬(SSRI)はうつ病性障害の第一選択薬である。セロトニンは選択的セロトニン再取り込み阻害薬(SSRI)の標的であり、脳内で利用可能なレベルを増加させる(Eicher and Mohajeri, 2022, Liang et al.) 腸脳軸とうつ病の関連性は示されているが、GMに対する抗うつ薬の効果を研究しているヒトの研究はわずかである(Macedo et al.) Shenらは、エスシタロプラムがGMを変化させ、Christensenellaceae、Eubacterium ruminantium group、Fusobacteriumが有意に増加し、Bacteroidesの有意な変化とともにLactobacillusの存在量がそれぞれ減少することを示した(Shenら、2021年)。双極性障害患者における非定型抗精神病薬治療の効果を研究したLe Bastardらの論文では、Lachnospiraceaeの相対存在量の増加(p= 0.029)、Akkermansia(p= 0.0006)とSutterellaの相対存在量の減少が示された(Flowersら、2017、Le Bastardら、2018)。オランダのコホートによる別の研究では、B. dorei(p= 0.051)とCoprococcus eutactus(p= 0.041)は抗うつ薬と正の相関を示し、Eubacterium halliiは負の相関を示した(p = 0.055)(Le Bastardら、2018)。
マウスモデルにおけるSSRI誘発性腸内変化に関する多くの研究が存在する(Sunら、2019、Zhangら、2021)。一般的な選択的セロトニン再取り込み阻害薬(SSRI)であるセルトラリン、フルオキセチン、パロキセチンは、ブドウ球菌や腸球菌などのグラム陽性菌に対して活性を示した。緑膿菌、クレブシエラ・ニューモニエ、シトロバクター属、M. morganii、クロストリジウム・ペルフリンゲンス、C. difficileなどの他の潜在的毒素原性腸内細菌も、SSRIの使用によってある程度減少した(Macedo et al.) これらの結果は、in vitro研究を根拠として見出された。フルオキセチンなどの特定の抗うつ薬を服用しているうつ病成人は、クロストリジウム・ディフィシル感染症(CDI)を発症しやすかった(Rogers et al.) マウスでは、エスシタロプラムとリチウムが同程度にセロトニン濃度を増加させることが報告されている(Bull-Larsen & Mohajeri, 2019)。しかしこの研究では、根本的な原因としてのディスバイオシスについては論じていない。うつ病に対するSSRIの作用機序は、これらの薬剤の抗菌作用とは無関係であるが、微生物群集の潜在的な変化は依然として見られ、気分に関連する他の炎症性パラメーターや生理的パラメーターに影響を及ぼす可能性がある(Flowers et al.)
考察
上述したように、多くの薬剤は意図した薬理作用に加えて、ディスバイオシスを誘発する可能性がある。したがって、他の生理学的経路や疾患もこれらの薬物によって影響を受けている可能性がある。ディスバイオーシスは多くの異なる疾患で観察されることから、マイクロバイオームの変化と疾患およびその治療との間に関連性があり、マイクロバイオームが疾患の発症に影響を及ぼす可能性が示唆される。そこで我々は、薬剤による腸内環境の変化が脳関連疾患にどのような影響を及ぼす可能性があるかに注目し、そのような疾患におけるマイクロバイオームの変化と比較した。以下の表では、薬剤誘発性腸内異常症と脳疾患に関連する腸内異常症の類似点が強調されている。次に、最も処方されている薬剤と疾患との関連性について議論した。
6.1. うつ病
うつ病性障害または大うつ病性障害は、最も一般的な精神神経疾患である。障害、罹患率、死亡率の主要な原因のひとつであり、生活の質を低下させる。およそ5人に1人が一生のうちに一度はうつ病と診断される。うつ病は複雑な疾患であり、症状も病態生理も様々である。DSM-5では、以下の症状が2週間以上持続する場合にうつ病と診断される:常に抑うつ気分、快感消失、孤独感、意欲低下、食欲・睡眠障害、精神運動興奮、集中困難、疲労、罪悪感や無価値感、自殺念慮(Amboss, 2023d)。病因や病態生理学は多くのメカニズムによって説明されるが、その全容はまだ明らかではなく、以下のようなものである: (1)セロトニン、ノルエピネフリン、ドーパミンのレベルが低く、グルタミン酸のレベルが高い(Troubat et al., 2021)、(2)HPA軸の変化、(3)免疫細胞、特にミクログリアへの誤った情報伝達につながる免疫メディエーターのアンバランスによる全身性の炎症、(4)そして最後に、MGBAによる微生物叢である。
現在までの研究で、うつ病患者の糞便では、門レベルではバクテロイデーテス門、プロテオバクテリア門、放線菌門、アクチノバクテリア門が増加し、一方、ファーミキューテス門が減少していることが判明している(Eicher and Mohajeri, 2022, Liang et al, 2022, Liang et al, 2018, Winter et al, 2018, Yao et al, 2023)。科・属レベルでは、腸内細菌科、エッゲルトヘラ属、ホルデマニア属、ゲリア属、ツリシバクター属、パラプレボテラ属、アネロフィラム属、ルミノコッカス科、連鎖球菌科、乳酸桿菌科、クロストリジウム属、ビフィズス菌科で増加が認められる。Bacteriodaceae、Lachnospiraceae、Prevotellaceae、Sutterellaceae、Veillonellaceaeでは減少が見られる(Changら、2022年、Eicher and Mohajeri、2022年、Knudsenら、2021年、Skonieczna-Żydeckaら、2018年、Yaoら、2023年、Zhaoら、2022年)。さらに属レベルでは、Faecalibacterium、Ruminococcus、Dialister、Oscillospiraceae、Bacteroides plebeius、Roseburia、Gemmiger、Parasutterella、Coprococcus、Escherichia/Shigellaが減少し、Desulfovibrioが増加している、 フラボニフラクター、アリスティペス、バクテロイデス、パラバクテロイデス、バルネシエラ、バクテロイデス・ヴァルガタス、アトポビウム、ワイセラ、ハロモナス、ブラウティア・クレブシエラ、アネロスチペス、クロストリジウム、ファスコラクトバクテリウム、ラクノスピラセ・インテルテセディス、ストレプトコッカス(Anand et al. , 2022、Changら、2022、Eicher and Mohajeri、2022、Knudsenら、2021、Liangら、2022、Liangら、2018、Skonieczna-Żydeckaら、2018、Valles-Colomerら、2019、Winterら、2018、Yaoら、2023)。Prevotellaceae、Prevotella、Bacteroides enterotype 2、BifidobacteriumおよびParasutterellaのような少数の細菌集団では、異なる研究において食い違いが認められた(Anand et al、 2022, Eicher and Mohajeri, 2022, Liang et al., 2018, Skonieczna-Żydecka et al., 2018, Valles-Colomer et al., 2019, Winter et al., 2018, Yao et al., 2023, Zhao et al., 2022)。
表5. メトホルミン、スタチン、PPI、NSAIDs使用者における細菌存在量の変化と関連したうつ病患者における細菌存在量の変化(それぞれの出典付き)。うつ病患者において存在量が多い(↑、緑色)または少ない(↓、オレンジ色)細菌。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意である。両方の矢印(↑↓)が描かれた灰色で着色されたセルは、より高い存在量とより低い存在量の両方で有意な変化があった。矢印のついていないセルは、相対的な変化のみを示し、有意差はない。
左の4行では、メトホルミン(MET)、スタチン、プロトンポンプ阻害薬(PPI)、非ステロイド性抗炎症薬(NSAIDs)による腸内細菌の変化を、うつ病患者に見られる腸内細菌集団の変化と比較している。
画像5
画像6
うつ病患者のGMと前述の薬剤誘発性腸内細菌異常症を比較すると、1つまたは複数の薬剤による異なる細菌分類レベルの類似性が観察される。バクテロイデス類はスタチンや非ステロイド系抗炎症薬とうつ病患者の腸内で増加している。バクテロイデスはグラム陰性のGABA産生菌で、免疫を誘発するLPSと抗炎症性のSCFAを持ち(Eicher and Mohajeri, 2022, Knuesel and Mohajeri, 2021)、免疫反応を引き起こす。うつ病は、セロトニンの枯渇(Knuesel & Mohajeri, 2021)を伴う全身性および神経炎症(Eicher & Mohajeri, 2022)と関連していることが示されている。しかし、バクテロイデス菌が産生するSCFAsとGABAを増加させれば、うつ病患者にはその量が少ないため、抑うつ行動の緩和に役立つと考えられる。SCFAを産生するルミノコッカス(Eicher and Mohajeri, 2022, Knuesel and Mohajeri, 2021)は、スタチンやPPIで増加し、SCFAの量の少なさを打ち消している。後者はまた、スタチンやPPIの経口投与後のうつ病患者におけるフェカリバクテリウムの存在量が少ない原因である可能性もある。フェカリバクテリウムは酪酸産生菌として知られており、腸管バリアの恒常性維持に重要であり、抗炎症種でもある(Eicher and Mohajeri, 2022, Knuesel and Mohajeri, 2021, Tran and Mohajeri, 2021, Valles-Colomer et al.) さらに、特にスタチンは、バクテロイデスの増加とフェーカリバクテリウムの減少の比率から生じるバクテロイデス腸型2(Bact2)を減少させることが示されている(Vieira-Silvaら、2020)。その結果、抑うつ症状が引き起こされる可能性がある(Eicher and Mohajeri, 2022, Knuesel and Mohajeri, 2021)。うつ病患者ではルミノコッカス数が少なく、スタチンやPPI使用者では多いことから、うつ病はスタチンやPPIの使用と負の相関がある可能性がある。クロストリジウムはメトホルミンとPPIでは減少するが、うつ病では減少しないことから、メトホルミンとPPIはクロストリジウムの減少に役立つ可能性がある。例えば、C. diffcileは有毒で感染性があるが、ClostridiumクラスターXIVaは有益なSCFAを産生する(Clooney et al.) セロトニンは迷走神経を介して体液性腸-脳経路に影響を与える可能性がある(González-Arancibia et al.) セロトニン合成には、腸内細菌科のエシェリヒア属と連鎖球菌が関与している(Eicher & Mohajeri, 2022)。これらの細菌は、うつ病患者の腸内にそれぞれ低濃度と高濃度で存在する。メトホルミンを使用するとエシェリヒア菌の増加が観察されることから、メトホルミンはエシェリヒア菌の低レベルを緩和する可能性がある。さらに、NSAIDsとPPIは、それぞれ腸内細菌科と連鎖球菌科で同じ効果を誘導する可能性があり、一方、スタチンは連鎖球菌の減少に正の相関を示し、PPIの効果を打ち消す可能性がある。バクテロイデス、ラクトバチルス、ビフィドバクテリウム、ストレプトコッカスもグルタミン酸経路に関与している可能性がある。したがって、これらの薬物誘発性の影響は、グルタミン酸作動性神経伝達を単独で、あるいは相互作用で上昇させる役割を果たす可能性がある(Eicher and Mohajeri, 2022, McGuinness et al.)
したがって、メトホルミン、スタチン、PPI、非ステロイド性抗炎症薬が誘発するディスバイオーシスは、それぞれうつ病の発症と進行に影響を及ぼす可能性があり、それらの作用は互いに打ち消し合ったり、強め合ったりする可能性がある。
6.2. 多発性硬化症
多発性硬化症はCNSの自己免疫疾患の中で最も多い疾患である。世界中で250万人以上が罹患しており、そのほとんどが若年成人である。多発性硬化症は、神経線維の脱髄と軸索の二次的破壊を特徴とする慢性の炎症性疾患であり、白質の組織病変によって特徴づけられる。症状は罹患した組織部位によって異なる。まず最も一般的な症状は、視神経炎による視覚障害と感覚障害および疲労である。間欠的な症状として、運動障害、疼痛、認知障害、心理的障害、植物障害があり、進行性または再発性である。炎症過程は、自己反応性末梢Tリンパ球が中枢神経系に侵入することで説明される(Amboss, 2023b)。しかし、この自己免疫反応の発生は不明である。遺伝的、アディポカインとサイトカインレベルの不均衡(Bonnechère, 2022)、エプスタイン・バー・ウイルス感染による自己反応性、ビタミンD欠乏、喫煙、MGBAを変化させる細菌やウイルスによる腸内細菌異常症などが、MSの病態生理に関与していると言われている(Thirion et al.、2023)。
MS患者は健常対照群と比べて微生物叢が乖離していることが示されている。MS患者の糞便を移植したGFマウスは、動物に見られる脱髄疾患である自己免疫性脳脊髄炎に共鳴する表現型を再現した。これらの結果は、健常対照の糞便を移植したGFマウスとは対照的であった(Doroszkiewicz et al.) 疾患の重症度や活動性さえも、異なる微生物組成と関連しているようである(Thirionら、2023)。一般に、抗炎症性の種の存在量が少ないことが観察されている(Tyler Patterson & Grandhi, 2020)。Thirionらは、未治療の患者において、臨床的に活動的な症例(すなわち、再発エピソードを有する症例)は、臨床的に活動的でない症例と比較してマイクロバイオームが豊富であることを発見した。細菌種を比較したところ、臨床的に活動的でないMS患者は、臨床的に活動的な患者に比べて、Faecalibacterium prausnitziiとGordonibacter urolithinfaciensの存在量が有意に高かった。同じ研究で、微生物組成が異なる61種が得られた。MSに濃縮された種は以下の通りである: Ruminococcus torques、Dysosmobacter welbionis、Flavonifractor plautii、Lawsonibacter phoceensis、Hungatella effluvia、Bilophila wadsworthia、Gordonibacter urolithinfaciens、Anaerobutyricum hallii、Pseudoflavonifractor capillosus、 Blautia wexlerae、Blautia massiliensis、Anaerotruncus colihominis、Erysipelatoclostridium ramosum、Ruminococcus gnavus、Sellimonas intestinalis、Coprobacillus cateniformis、Clostridium innocuum。健常対照と比較して減少した細菌種は、Haemophilus parainfluenzae、Veillonella rogosae、Victivallis vadensis、Bifidobacterium angulatum、Streptococcus australisなどであった(Thirionら、2023)。全体として、MS患者は健常対照者よりも微生物組成の豊かさが低かった。これはまた、MS関連細菌種と相関する特異的炎症性バイオマーカーの存在量の高さとも関連していた(Thirion et al.)
他の研究でも同様の結果が得られている:アクチノミセス属、アッカーマンシア属、ビフィオデバクテリウム属、コプロコッカス属、ダイアリスター属、ドレア属、ヘモフィルス属、メガファエラ属、パラプレボテラ属、シュードモナス属、マイコプラナ属、ブラウチア属、ルミノコッカス属、ストレプトコッカス属の有意な増加(Bonnechère, 2022, Bonnechère et al、 2022、Chenら、2021、Doroszkiewiczら、2021、Erturk-Hasdemirら、2021、Pellegriniら、 2018, Tran and Mohajeri, 2021, Tyler Patterson and Grandhi, 2020);ブチリコッカス(Butyricicoccus)、ゲンミガー(Gemmiger)、パラバクテロイデス(Parabacteroides)、ファスコアクトバクテリウム(Phascoarctobacterium)、プレボテラ(Prevotella)、コリンセラ(Collinsella)、ラクトバチルス(Lactobacillus)、アドレルクロイツィア(Adlercreutzia)、アネアロスティペス(Anearostipes)の減少(Bell et al、 2019, Bonnechère, 2022, Bonnechère et al., 2022, Chen et al., 2021, Doroszkiewicz et al., 2021, Erturk-Hasdemir et al., 2021, Tran and Mohajeri, 2021, Tyler Patterson and Grandhi, 2020)。全体として、バクテロイデス門の減少が見られ(Chenら、2021年、Doroszkiewiczら、2021年、Erturk-Hasdemirら、2021年、Pellegriniら、2018年)、B. coprocola、B. coprophilus、B. stercorisなどの種がそれぞれ減少した(Pellegriniら、2018年)。Faecalibacterium、Slackia、Clostridium、Methanobrevibacter、Butyricimonasでは一貫性のない結果が見られた(Bellら、2019、Bonnechère、2022、Chenら、2021、Doroszkiewiczら、2021、Erturk-Hasdemirら、2021、Tyler Patterson and Grandhi、2020)。
アッカーマンシア(Akkermansia)の増加とプレボテラ(Prevotella)の減少は、MSに関するすべての引用文献において、一貫して変化した細菌集団のひとつであった。
表6. メトホルミン、スタチン、PPIおよびNSAIDs使用者における細菌量の変化と関連した、多発性硬化症(MS)患者における細菌量の変化(それぞれの由来とともに)。うつ病患者において存在量が多い(↑、緑色)、または少ない(↓、オレンジ色)細菌。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意である。両方の矢印(↑↓)が描かれた灰色で着色されたセルは、より高い存在量とより低い存在量の両方で有意な変化があった。矢印のついていないセルは、相対的な変化のみを示し、有意差はない。左の4行では、メトホルミン(MET)、スタチン、プロトンポンプ阻害薬(PPI)、非ステロイド性抗炎症薬(NSAIDs)による腸内細菌の変化を、多発性硬化症患者に見られる腸内細菌集団の変化と比較している。
画像7
画像8
グラム陰性菌属でSCFAやGABAを産生するバクテロイデス(Eicher & Mohajeri, 2022)は、MS患者とは対照的にスタチンやNSAIDs使用者で増加している。別のグラム陰性菌種であるBilophila wadsworthia(Eicher & Mohajeri, 2022)は、MSとMETでは増加するが、スタチンでは減少することから、MS患者にとってMETは不利でスタチンは有利である可能性が示唆される。グラム陰性菌が宿主の免疫反応を誘導するのは、その細胞壁にLPSが存在するからである。レンサ球菌属も同様で、MS患者やPPI使用者では増加し、スタチン使用者では減少する。日和見菌であるレンサ球菌は、神経毒である酢酸やセロトニン(5-HT)を産生することがある(Eicher & Mohajeri, 2022)。一方、ブラウチア属はMS、MET、PPIで増加する。ブラウチア属は酪酸産生菌であり(Eicher and Mohajeri, 2022, Tong et al., 2018, Tran and Mohajeri, 2021)、腸やBBBなどのバリア統合性やミクログリアの成熟・活性化に重要である(Eicher & Mohajeri, 2022)。クロストリジウムはMETやPPIの使用者では減少しているが、多発性硬化症患者では曖昧な存在量である。クロストリジウムはSCFAを産生し、Treg細胞に影響を与える可能性がある(Doroszkiewiczら、2021、Tran and Mohajeri、2021)。SCFA産生菌であるルミノコッカス(Ruminococcus)は、MSやスタチンやPPI使用後に増加する。LPS産生菌でGABA産生菌(Eicher & Mohajeri, 2022)やプロピオン酸産生菌(Chen et al., 2021, Doroszkiewicz et al., 2021)であるPrevotellaは、NSAIDsで増加し、MSやPPI使用後では減少する。BifidobacteriumとLactobacillusは、GABAと酢酸の生産者(Eicher & Mohajeri, 2022)であり、それぞれMSとMET摂取後に増加、減少しており、この2つの属がMSの疾患と進行に影響を及ぼす可能性が示唆された。
この結果を見ると、MSと薬剤誘発性ディスバイオシスとの間には多くの関連性がある可能性がある。とはいえ、これらの仮説を証明するには、より明確な研究結果が必要である。
6.3. パーキンソン病
MGBAの分野では、パーキンソン病(PD)は最も研究が盛んな神経変性疾患である。主な症状は、アキネジー、硬直、安静時振戦、姿勢不安定などの運動症状で、主に黒質下部のドーパミン欠乏に起因する。レビー小体凝集体は、ミスフォールディングしたタンパク質の凝集体、すなわちアミロイド(主にα-シヌクレインアミロイド)からなり、ドーパミン作動性ニューロンの破壊の主な原因である(Amboss, 2023c)。しかし、このような凝集体がどのようにして生成されるのか、詳しいことはまだわかっていない。
PD患者は非運動症状に大きく影響され、その多くは運動障害が発症する数十年前に先行する。胃腸機能障害、抑うつ気分、睡眠障害、筋肉痛、関節痛はこれらの前駆症状の一部である(Amboss, 2023c)。前者では、閉塞や胃排出遅延、唾液漏、嚥下障害、胃下垂、SIBOが最もよくみられる(Ryman et al., 2023)。これらの前駆期胃腸障害を有する患者は、PDの進行がより重篤である(Rymanら、2023年)。アミロイドの形成はENSニューロン、特にマウスやPD患者のPD初期にも観察されている(Wittung-Stafshede, 2022)。最近の研究では、マウスの消化管内のアミロイド線維によって脳内のレビー小体形成が誘導され、三叉迷走神経切断術によってそれらのレビー小体形成のリスクが劇的に減少することが示された。
アミロイド形成は、一つのタンパク質だけに特異的である必要はない。アミロイド形成蛋白質は互いに交雑してアミロイドを形成することがある。その分子過程はまだ解明されていないが、ヒト同士、あるいはヒトとヒト以外のアミロイド形成タンパク質の間で起こりうる。GMにはアミロイドを含むバイオフィルムを産生する種がいくつかある。腸アミロイドは理論上、迷走神経経路を通って脳へ移動し、αシヌクレインを交差播種する可能性が示唆されている(Pellegrini et al.) 大腸菌、シュードモナス、連鎖球菌、ブドウ球菌、サルモネラ、マイコバクテリア、クレブシエラ、シトロバクター、バチルスが、このような細胞外アミロイドを産生する腸内細菌の候補である(Wittung-Stafshede, 2022)。Nuzumらも、SCFA産生菌の存在量が全体的に低いことを示唆している(Nuzumら、2020)。従って、腸内細菌叢はPDの発症を引き起こすだけでなく、調節する可能性もある。
PD患者と健常対照者の微生物組成を比較すると、ルミノコッカス科、疣贅菌科、ポルフィロモナス科、アッカーマンシア科(A. muciniphila種を含む)、Pasteurellaceae、Lachnospiraceae、Desulfovibrio、Lactobacillus、Bifidobacterium(B. adolescentis種を含む)、Ralstonia、Enterobacteriaceae、Flavonifractor、Cornebacterium、Alistipes、EscherichiaおよびMegasphaeraがPD患者で優勢であることが報告されている(Bell et al、 2019、Chapeletら、2019、Chen and Lin、2022、Gerhardt and Mohajeri、2018、Liら、2019、Nuzumら、2020、Pellegriniら、2018、Rajputら、2021、Rymanら、2023、Zhangら、2022)。Bacteroidetes、Prevotella、Faecalibacterium(F. prausnitziiを含む)、Coprococcus、Blautia、Roseburia(R. faecisを含む)、Eubacterium、Anaerostipes、Veillonella parvula、EnterococcaceaeおよびDoreaは、枯渇した存在量を示すことが判明した(Bell et al、 2019, Chapelet et al., 2019, Chen and Lin, 2022, Gerhardt and Mohajeri, 2018, Li et al., 2019, Nuzum et al., 2020, Pellegrini et al., 2018, Rajput et al., 2021, Ryman et al., 2023, Zhang et al., 2022)。乳酸桿菌科は、高い存在量と低い存在量の両方を示し、一貫して観察されなかった。
表7. メトホルミン、スタチン、PPIおよびNSAIDs使用者における細菌存在量の変化と関連したパーキンソン病(PD)患者における細菌存在量の変化(それぞれの由来とともに)。うつ病患者において存在量が多い(↑、緑色)または少ない(↓、オレンジ色)細菌。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意である。両方の矢印(↑↓)が描かれた灰色で着色されたセルは、より高い存在量とより低い存在量の両方で有意な変化があった。矢印のついていないセルは、相対的な変化のみを示し、有意差はない。左の4行では、メトホルミン(MET)、スタチン、プロトンポンプ阻害薬(PPI)、非ステロイド性抗炎症薬(NSAIDs)による腸内細菌の変化を、パーキンソン病患者に見られる腸内細菌集団の変化と比較している。
画像9
画像10
パーキンソン病の腸内細菌叢異常と薬剤性腸内細菌叢異常を比較すると、以下のようになる: PPIはSIBOを誘発し(Kooら、2019)、サルモネラ菌感染のリスクを高める(Naitoら、2018)。SIBOはPD患者にも認められ(Rymanら、2023)、サルモネラは細胞外アミロイド産生因子である(Wittung-Stafshede、2022)ことから、両者ともPDに影響を及ぼす可能性がある。しかし、サルモネラはPPIとPDの共通項としては観察されない。MSと同様に、ルミノコッカス科はPD、スタチン、NSAIDsで増加している。しかし、SCFAはBBBの完全性を強化することが知られており(Eicher & Mohajeri, 2022)、その結果、炎症性因子が脳を通過するリスクが減少し、細胞外アミロイドの形成が抑制される。一方、ファエカリバクテリウム(Faecalibacterium)は、腸とBBBの完全性に重要な酪酸産生細菌であり、抗炎症性細菌群として知られている(Eicher and Mohajeri, 2022, Tran and Mohajeri, 2021)が、PD、スタチン、PPIでは減少していることから、ファエカリバクテリウムがパーキンソン病患者におけるバリアの弱体化と炎症プロセスの亢進の原因の一つである可能性が示唆される。しかしながら、METとPPIはFaecalibacteriumの不利な点を覆すのに役立つかもしれない。というのも、METとPPIはBlautiaの存在を増加させるが、PD患者では減少するからである。同様に、スタチンや非ステロイド性抗炎症薬によってルミノコッカス科の存在量が増加することも、フェカリバクテリウムや他の不利な菌種の影響を補う可能性がある。例えば、PPIはBlautia(有利なSCFA産生者(Eicher & Mohajeri, 2022; Ryman et al., 2023; Tran & Mohajeri, 2021))を増加させ、同時にSIBO(Koo et al.
全体として、METの摂取は、この論文で言及した他の薬剤よりも、パーキンソン病で見られる効果と最も類似した効果をマイクロバイオームにもたらす。Akk.muciniphila(SCFA産生およびLPS産生菌(Eicher & Mohajeri, 2022; Tran & Mohajeri, 2021))、B. adolescentis(GABA産生菌(Eicher & Mohajeri, 2022))、Escherichia(LPS産生菌、5-HT代謝菌(Eicher & Mohajeri, 2022))がPD患者とMET使用者の両方で観察され、このうち前者と後者の菌種は細胞壁にLPSを持つグラム陰性菌であるが、それぞれSCFAと5-HTも産生する。繰り返しになるが、両者とも有利な作用と不利な作用を持つことから、ある疾患の病因の主要な決定因子は1つの細菌ではないことが示唆される。むしろ、疾患の発症と進行に正味の影響を及ぼすには、それらの相互作用と相対的な存在量が重要であるようだ。
6.4. アルツハイマー病
アルツハイマー病(AD)は老年痴呆の最も一般的な形態であり、全世界の痴呆症例の約50-70%を占めている。その有病率は年齢とともに増加し、世界の人口が高齢化していることから、ADの発生率は今後数年でより高くなると考えられている。ほとんどの場合、新しく形成された記憶の喪失から始まり、言語や視空間指向性などの神経認知機能や実行機能がさらに低下する(Amboss, 2023a)。
病態生理学は多面的であるが不明であり、不可逆的な神経細胞やシナプスの喪失につながる。加齢に加え、不溶性のアミロイドβ斑の蓄積、それに続くタウタンパク質の神経原線維のもつれ、神経伝達物質の不均衡、神経炎症などがADの特徴である(Amboss, 2023a)。腸内細菌叢は加齢とともに大きく変化するため、加齢に伴う腸内細菌叢異常がADの病因にも関与している可能性が示唆される。高齢者では、炎症性細菌と抗炎症性細菌の比率が増加し、SCFA産生細菌が減少することで、腸管リーキー、BBBの障害、ミクログリアの活性化につながり、その豊かさの低下と組成の変化がより顕著になる(Eicher & Mohajeri, 2022)。ミクログリアはアベータ斑を除去するのに重要であり、その能力がアベータの産生増加や炎症性構造変化による能力低下によって妨げられると、アミロイド斑がアルツハイマー病患者の脳に蓄積する可能性がある(Eicher and Mohajeri, 2022, Zhou et al.) アミロイドタンパク質は細菌由来であると推測されており、PDと同様に、脳内の他のアミロイド構造を交差播種するか、リーキーガットによる全身性炎症から神経炎症を悪化させる可能性がある(Eicher & Mohajeri, 2022)。さらに、SIBO呼気試験が有意に陽性(p=0.025)であったことによる細菌の転移していることも報告されている(Kowalski & Mulak, 2022)。AD患者における細菌の移動とLPSレベルの増加は、ともに神経炎症を高める(D'Argenio and Sarnataro, 2019, Doroszkiewicz et al., 2021, Epishina and Budanova, 2022)。
主な知見は、炎症性細菌Escherichia/Shigellaの増加と抗炎症性細菌Eubacterium rectale種の減少を指摘している(Chenら、2021年、Choiら、2022年、D'Argenio and Sarnataro、2019年、Doroszkiewiczら、2021年、Wuら、2022年)。プロ炎症性サイトカインと抗炎症性サイトカインの間にそれぞれの相関関係が見つかっており(Choiら、2022)、また大腸菌はアミロイド産生菌に属する(Choiら、2022、Doroszkiewiczら、2021)ことから、これらのアンバランスは認知障害とアミロイド合成の主な要因の一つである可能性がある。
AD患者では、バクテロイデーテス(Bacteroidetes)属、ルミノコッカス(Ruminococcaceae)属(Chenら、2021、Zhouら、2022)、プレボテラ(Prevotellaceae)属、エンテロコッカス(Entereococcaceae)属、ラクトバチルス(Lactobacillaceae)属、ドレア(Dorea)属、ストレプトコッカス(Streptococcus)属(Chenら、2021)の増加が報告された。一方、ビフィズス菌、Bacillus/Bacteroides fragilis、Eubacterium hallii、Faecalibacterium prausnitzii(Eicher & Mohajeri, 2022)、Fusobacteriacaea、Bacteroides fragilis、NegativicutesおよびVeillonellaceae(Chen et al. Firmicutes(Eicher and Mohajeri, 2022, Zhou et al., 2022)、Lactobacillus(Chen et al., 2021, D'Argenio and Sarnataro, 2019, Zhou et al., 2022)、LachnospiraceaeaおよびBacteroidaceae(Chen et al., 2021, Zhou et al., 2022)では一貫性のない結果が得られた。
表8. メトホルミン、スタチン、PPI、非ステロイド性抗炎症薬(NSAIDs)使用者における細菌量の変化と関連したアルツハイマー病(AD)患者における細菌量の変化(それぞれの供給源とともに)。うつ病患者において存在量が多い(↑、緑色)、または少ない(↓、オレンジ色)細菌。色のついたセルは、少なくともp<0.05の有意水準を表し、言及された情報源の少なくとも1つにおいて有意である。両方の矢印(↑↓)が描かれた灰色で着色されたセルは、より高い存在量とより低い存在量の両方で有意な変化があった。矢印のついていないセルは、相対的な変化のみを示し、有意差はない。左の4行では、メトホルミン(MET)、スタチン、プロトンポンプ阻害薬(PPI)、非ステロイド性抗炎症薬(NSAIDs)による腸内細菌の変化を、アルツハイマー病患者に見られる腸内細菌集団の変化と比較している。
画像11
上記の薬剤とアルツハイマー病患者のうち、スタチン使用との関連は、微生物叢の変化と最も強い関連を示した。特に、酪酸産生菌であるFaecalibacterium prausnitzii(Eicher and Mohajeri, 2022, Knuesel and Mohajeri, 2021)がスタチン使用者で増加しており、酪酸が抗炎症過程やミクログリアの活性化と成熟に大きな役割を果たしていることから、BBBの維持に役立っている可能性がある(Eicher & Mohajeri, 2022)。連鎖球菌は細胞外アミロイドを産生することが示唆されているため、PPIと併用するスタチンは、PPIの使用による連鎖球菌の増加を相殺するのに役立つかもしれない。SCFAを産生するルミノッカス科の細菌(Eicher & Mohajeri, 2022)は、スタチンやNSAIDの使用者、AD患者で増加しており、おそらくパーキンソン病(PD)と同様の代償作用があると考えられる。PDと同様に、AD患者でもSIBOの結果が陽性であることから、NSAIDsの経口投与の有無にかかわらず、PPIが大きな寄与をしている可能性がある。
結論と今後の展望
これまでの知見から、神経疾患に対する腸内細菌叢異常の影響は大きいことが示されている。微生物叢の多様性と豊かさはともに変化している。抗生物質以外の薬剤による変化をMGBAと組み合わせた研究はまだほとんどない。関連するヒトのデータと前臨床研究で発表されたメカニズムの裏付けとなるデータをすべて考慮に入れて、メトホルミン、スタチン、PPI、NSAIDsがどのように、そしてなぜMGBAを変化させるのかを、うつ病、多発性硬化症、パーキンソン病、アルツハイマー病を神経疾患の例として用いて概説した。われわれのデータは、薬物投与や神経細胞疾患によるディスバイオティクス環境において、一般的な細菌株が存在することを示している。ある疾患状況において、特定の条件下でディスバイオシスを緩和できる薬剤は少ないが、ディスバイオシスを生じさせる可能性もある。一方、有利な作用と不利な作用の両方を持つ薬剤もあった。有利なSCFA産生菌であるルミノコッカス属は、上記のすべての薬剤使用者および疾患において上昇している。プレボテラ(Prevotella)とアッカーマンシア(Akkermansia muciniphila)はいずれも細胞壁にLPSを持ち、有益なSCFAを産生する。エシェリヒア属もLPSキャリアーであり、5-HTを代謝し、細胞外アミロイドの前駆体となる。クロストリジウム属には、クロストリジウム・ディフィシル(Clostridium difficile)のような有毒種とSCFA産生種がある。うつ病ではSCFAやGABAが減少しているが、バクテロイデス属は増加している。もし病気がスタチンやNSAIDsの使用と相関していれば、バクテロイデス属は豊富に存在することになる。バクテロイデス属はGABAとSCFAを産生し、細胞壁にLPSを持つ。これらの例から、特定の細菌株ではなく、細菌集団の相対的な存在量とそれらの相互作用がMGBAに影響を及ぼすことが示唆される。さらに、取り上げた薬剤は、合併症のある患者において併用されることが多い。従って、これらの薬剤の作用は互いに相殺し合うか、あるいは細菌集団のそれぞれの変化を強め、神経疾患におけるディスバイオーシスと正または負に関連する可能性がある。一般に、研究の診断方法、腸内細菌叢または糞便微生物叢の検出方法、患者の除外/包含基準などが、実施された研究のほとんどでかなり異なるため(例えば、(Knudsen et al.) 研究によって、細菌検体は上部、中部、下部消化管、または便検体のいずれかであった。本レビューで取り上げたすべての論文が、起こりうる交絡因子(原疾患の状態、他の薬剤の併用、薬剤投与量、薬剤曝露量、薬剤摂取期間、年齢、性別、患者の食生活、併存疾患、民族性など)をすべて考慮に入れているわけではなく、また適切な対照群やコホートサイズを欠いているわけでもなかった。さらに、対照群は健康な患者ではなく、すでに疾患と診断された患者であった。いずれの場合も、それぞれの対照コホートとの比較について言及している。さらに、著者らは臨床研究の結果をさらに裏付けるために、ヒトの糞便細菌サンプルを抗生物質処理マウス(無菌マウス)に移植した。全体として、このような制約があるため、結果に一貫性がない可能性がある。
明確ではないにせよ、薬物使用とMGBAの関連、MGBAと神経疾患の関連は、それぞれ強い証拠によって示唆されている。今後の研究では、体性疾患の治療に使用される医薬品と、その腸内環境を変化させるMGBAが、脳関連疾患の発症や進行に及ぼす影響に焦点を当てることができるだろう。腸内細菌叢は様々な環境要因や生活習慣要因によって変化するため、このような研究では併存する高齢者も考慮すべきである。複数の薬剤を服用していたり、食欲不振で食生活が乱れていたりすると、高齢者の腸内細菌叢にはさまざまな変化が生じることが予想される。しかし、日常的に処方される薬剤によって誘発される腸内細菌叢異常と、MGBAを介した脳関連疾患への影響との相関関係を探ることは、まだ新しいテーマである。これらの相関関係についての十分なデータは、示唆に富む結論を導き出すには存在しない。この知識のギャップを埋めることは、MBGAの双方向コミュニケーションをよりよく理解し、新規治療法を用いて患者を個別に治療する上で、新たな重要な展望をもたらすかもしれない。
引用されていない文献
(de la Cuesta-Zuluaga et al.
資金提供
本研究は、公的、営利、非営利のいずれの分野の助成機関からも特定の助成を受けていない。
CRediT著者貢献声明
Garg Kitri:執筆(原案)、視覚化、検証、ソフトウェア、方法論、形式分析、データキュレーション、概念化。モハジェリ・M・ハサン 執筆-校閲・編集、バリデーション、監修、プロジェクト管理、形式分析、概念化。
利益相反宣言
著者らは利益相反がないことを宣言する。
謝辞
David P. Wolfer、Prof. MD、Julinka Waesche、Yogendra Garg、Luzi Brunnerの有益なコメントと本論文の査読に感謝する。
付録A. 補足資料
ダウンロード ワード文書ダウンロード(4MB)
補足資料
データの入手
データはご要望に応じて提供いたします。
参考文献
NCBI, 2023
NCBI)、N.分類学ブラウザ。で。
グーグル・スカラー
アダックとカーン、2019
A. アダック、M.R.カーン
腸内細菌叢とその機能性に関する洞察
細胞分子生命科学, 76 (3) (2019), pp.473-493, 10.1007/s00018-018-2943-4
Scopusで見るGoogle Scholar
アギルマンら、2021年
G.アギルマン、K.B.ユー、E.Y.シャオ
腸-脳軸を横断するシグナル伝達炎症
Science, 374 (6571) (2021), 1087-1092頁, 10.1126/science.abi6087
スコープで見るGoogle Scholar
アグニホトリとモハジェリ、2022年
N. アグニホトリ、M.H.モハジェリ
成人の神経新生と脳由来神経栄養因子の発現における腸内細菌叢の関与
Int J Mol Sci, 23 (24) (2022), 10.3390/ijms232415934
グーグル奨学生
アルムガダムら, 2020
B.S.アルムガダム、Y.リウ、S.M.チェン、C.H.ワン、C.Y.シャオ、B.W.レン、L.タン
2型糖尿病患者における腸内細菌叢の変化と抗糖尿病薬の交絡効果
J Diabetes Res, 2020 (2020), Article 7253978, 10.1155/2020/7253978
スコープで見るGoogle Scholar
アンボス、2023a
アンボス (2023a). アルツハイマー病。〈https://next.amboss.com/de/article/D301kf?q=alzheimer-demenz#Z355d10c7e7f39ad060b2a6c28268f874〉。
グーグル・スカラー
アンボス、2023b
アンボス.(2023b). 複数のスクレーローズ。〈https://next.amboss.com/de/article/WR0PNf?q=multiple+sklerose#Zfc37457ca8f0aea91134b8e6e325e935〉。
グーグル・スカラー
アンボス, 2023c
Amboss. (2023c). パーキンソン症候群とパーキンソン病。〈https://next.amboss.com/de/article/C30qkf〉.
グーグル・スカラー
アンボス, 2023d
アンボス.(2023d). ユニポラーレうつ病。〈https://next.amboss.com/de/article/PP0WUT?q=unipolare+depression#Z992ea889338baceb63e016ee83bfc473〉。
Google Scholar
アナンドら、2022
N. アナンド、V.R.ゴラントラ、S.B.チダンバラム
精神神経疾患の病態生理における腸内細菌異常症の役割
Cells, 12 (1) (2022), 10.3390/cells12010054
グーグル奨学生
バジャジら、2018
J.S. Bajaj、C. Acharya、A. Fagan、M.B. White、E. Gavis、D.M. Heuman、P.M. Gillevet
プロトンポンプ阻害薬の投与開始と中止は肝硬変における腸内細菌叢と再入院リスクに影響する
Am J Gastroenterol, 113 (8) (2018), 1177-1186頁, 10.1038/s41395-018-0085-9
スコープで見るGoogle Scholar
ビームら、2021
A. ビーム、E.クリンガー、L.ハオ
腸内細菌叢の組成に及ぼす食事と食事成分の影響
栄養素, 13 (8) (2021), 10.3390/nu13082795
Google Scholar
ベルダとアルマリオ, 2009
X. ベルダ、A. アルマリオ
ドーパミンD1およびD2受容体は、不動ストレスによる視床下部-下垂体-副腎軸の活性化を制御する。
Psychopharmacology (Berl), 206 (3) (2009), pp.355-365, 10.1007/s00213-009-1613-5
Scopusで見るGoogle Scholar
ベルら、2019年
J.S.ベル、J.I.スペンサー、R.L.イェーツ、S.A.イー、B.M.ジェイコブス、G.C.デルーカ
招待総説: 鼻から腸まで-神経疾患におけるマイクロバイオームの役割
Neuropathol Appl Neurobiol, 45 (3) (2019), pp.195-215, 10.1111/nan.12520
ScopusGoogle Scholarで見る
ビンドゥ・シャジャン・ペラッパダン, 2015
ビンドゥ・シャジャン・ペラッパダン、T.H. (2015). 52%のインド人がセルフメディケーションに耽っている:調査。〈https://www.thehindu.com/news/cities/Delhi/52-per-cent-indians-indulge-in-selfmedication-survey/article7096902.ece〉.
グーグル・スカラー
ボナズら、2018
B. ボナズ、T.バザン、S.ペリシエ
微生物叢-腸-脳軸の界面における迷走神経
Front Neurosci, 12 (2018), Article 49, 10.3389/fnins.2018.00049
Scopusで見るGoogle Scholar
ボンヌシェール、2022
B. ボンヌシェール
多発性硬化症の管理におけるマルチオミクスアプローチへのリハビロミクスの統合: 精密医療への道?
Genes (Basel), 14 (1) (2022), 10.3390/genes14010063
Google Scholar
Bonnechère et al.
B. ボンヌシェール、N.アミン、C.ファン・ドゥイン
神経疾患に関与する主要な腸内細菌叢とは何か?系統的レビュー
Int J Mol Sci, 23 (22) (2022), 10.3390/ijms232213665
グーグル・スカラー
ブリラップら、2019
T. Bryrup, C.W. Thomsen, T. Kern, K.H. Allin, I. Brandslund, N.R. Jørgensen, T. Nielsen
健康な若年男性におけるメトホルミンによる腸内細菌叢の変化:非盲検単群介入試験の結果
Diabetologia, 62 (6) (2019), 1024-1035頁, 10.1007/s00125-019-4848-7
グーグル・スカラー
ブル=ラーセン、モハジェリ、2019年
S. ブル=ラーセン、M.H.モハジェリ
ADHDの発症と進行に対する細菌マイクロバイオームの潜在的影響
栄養素, 11 (11) (2019), 10.3390/nu11112805
グーグル・スカラー
チャクラバルティら、2022年
A. チャクラバルティ、L.グールツ、L.ホイルズ、P.イオッツォ、A.D.クラネヴェルト、G.ラファタ、D.ヴォズール
微生物叢-腸-脳軸:より良い脳の健康への道。知っていること、調査すべきこと、知識を実践する方法についての視点
Cell Mol Life Sci, 79 (2) (2022), Article 80, 10.1007/s00018-021-04060-w
グーグル奨学生
チャンら、2022年
L. Chang, Y. Wei, K. Hashimoto
うつ病における脳-腸-微生物叢軸: 歴史的概観と今後の方向性
Brain Res Bull, 182 (2022), 44-56頁, 10.1016/j.brainresbull.2022.02.004
PDFを見る記事を見るScopusGoogle Scholarで見る
シャペレら、2019
G.シャプレ、L.ルクレール=ヴィゾノー、T.クレアムボー、M.ノインリスト、P.ダーキンデレン
腸は、PD病態を解明する上で欠落したピースとなり得るか?
Parkinsonism Relat Disord, 59 (2019), pp.26-31, 10.1016/j.parkreldis.2018.11.014
PDFで記事を見るScopusで記事を見るGoogle Scholar
チェンとリン、2022年
S.J.チェン、C.H.リン
腸脳軸を介したパーキンソン病の引き金となりうる腸内微小環境の変化
J Biomed Sci, 29 (1) (2022), Article 54, 10.1186/s12929-022-00839-6
スコープで見るGoogle Scholar
チェンら、2021年
Z. チェン、J.マクブール、F.サジド、G.フサイン、T.サン
ヒト腸内細菌叢と神経変性疾患の病因および治療との関連性
Microb Pathog, 150 (2021), Article 104675, 10.1016/j.micpath.2020.104675
PDFで記事を見るScopusで記事を見るGoogle Scholar
Choi et al.
H. Choi, D. Lee, I. Mook-Jung
アルツハイマー病の病態形成における隠れたプレーヤーとしての腸内細菌叢
J Alzheimers Dis, 86 (4) (2022), 1501-1526頁, 10.3233/jad-215235
スコープで見るGoogle Scholar
ClinCalc.com, 2020
ClinCalc.(2020). 2020年のトップ300医薬品。で。
Google Scholar
クルーニーら、2016
A.G.クルーニー、C.N.バーンスタイン、W.D.レスリー、K.ヴァギアノス、M.サージェント、E.J.レーザーナ-メンディエタ、L.E.ターゴーニック
プロトンポンプ阻害薬の長期使用者と非使用者の腸内細菌叢の比較
Aliment Pharmacol Ther, 43 (9) (2016), pp.974-984, 10.1111/apt.13568
Scopusで見るGoogle Scholar
クライアンら、2019
J.F.クライアン、K.J.オリオルダン、C.S.M.コーワン、K.V.サンドゥ、T.F.S.バスティアンスセン、M.ベーメ、T.G.ディナン
微生物叢-腸-脳軸
Physiol Rev, 99 (4) (2019), 1877-2013頁, 10.1152/physrev.00018.2018
Scopusで見るGoogle Scholar
クッソットら、2018
S. クッソット、K.V.サンドゥ、T.G.ディナン、J.F.クライアン
微生物叢-腸-脳軸の神経内分泌学:行動学的視点
Front Neuroendocrinol, 51 (2018), 80-101頁, 10.1016/j.yfrne.2018.04.002
PDFで記事を見るScopusで記事を見るGoogle Scholar
ダルゲニオとサルナータロ、2019年
V. ダルゲニオ、D.サルナータロ
プリオン病とアルツハイマー病の発症におけるマイクロバイオームの影響
Int J Mol Sci, 20 (19) (2019), 10.3390/ijms20194704
グーグル・スカラー
ダリレら、2019
B. ダリレ、L.ヴァン・オーデンホーフ、B.ヴァーヴリート、K.フェルベーク
微生物叢-腸-脳のコミュニケーションにおける短鎖脂肪酸の役割
Nature Reviews Gastroenterology & Hepatology, 16 (8) (2019), pp.461-478, 10.1038/s41575-019-0157-3
Scopusで見るGoogle Scholar
デ・ラ・クエスタ=ズルアガら、2017年
J. de la Cuesta-Zuluaga、N.T. Mueller、V. Corrales-Agudelo、E.P. Velásquez-Mejía、J.A. Carmona、J.M. Abad、J.S. Escobar
メトホルミンは、腸内のムチン分解性Akkermansia muciniphilaおよびいくつかの短鎖脂肪酸産生微生物叢の相対存在量の増加と関連している
糖尿病ケア, 40 (1) (2017), 54-62頁, 10.2337/dc16-1324
スコープで見るGoogle Scholar
Dengら、2022年
X. Deng、C. Zhang、P. Wang、W. Wei、X. Shi、J. Yang、H. Yuan
エンパグリフロジンの心血管ベネフィットは2型糖尿病における腸内細菌叢と血漿代謝物に関連する
J Clin Endocrinol Metab, 107 (7) (2022), pp.1888-1896, 10.1210/clinem/dgac210
スコープで見るGoogle Scholar
Doroszkiewiczら、2021年
J. ドロシュキエヴィチ、M.グロブレフスカ、B.ムロツコ
中枢神経系の選択された疾患における腸内細菌叢と腸-脳の相互作用の役割
Int J Mol Sci, 22 (18) (2021), 10.3390/ijms221810028
Google Scholar
アイヒャーとモハジェリ, 2022
T.P.アイヒャー、M.H.モハジェリ
一般的な脳疾患の病態形成における脳内活性細菌および細菌代謝産物の作用機序の重複
栄養素, 14 (13) (2022), 10.3390/nu14132661
グーグル奨学生
エピシナおよびブダノワ, 2022
I.V.エピシナ、E.V.ブダノワ
[神経変性疾患の発症におけるヒト微生物叢の役割]
神経変性疾患の発症におけるヒト微生物叢の役割] Zh Nevrol Psikhiatr Im S S Korsakova, 122 (10) (2022), pp.57-65, 10.17116/jnevro202212210157
(滋養強壮のための微生物療法, 122 (10) (2022), pp.10.176/jnevro2022122157.
スコープで見るGoogle Scholar
Erturk-Hasdemirら、2021年
D. Erturk-Hasdemir, J. Ochoa-Repáraz, D.L. Kasper, L.H. Kasper
中枢神経系炎症性脱髄の制御における腸脳軸の探索: バクテロイデス・フラジリスの多糖体Aによる免疫調節作用
Front Immunol, 12 (2021), Article 662807, 10.3389/fimmu.2021.662807
スコープで見るGoogle Scholar
エスマットら、2023年
N. エスマット、D.Á. Karádi, F. Zádor, K. Király, S. Fürst, M. Al-Khrasani
オピオイド誘発性便秘とディスバイオシスに関するオピオイド拮抗薬の現在および将来的使用可能性に関する洞察
分子, 28 (23) (2023)
Google Scholar
フラワーズら、2017年
S.A.フラワーズ、S.J.エヴァンス、K.M.ウォード、M.G.マクニス、V.L.エリングロッド
双極性障害コホートにおける非定型抗精神病薬と腸内細菌叢の相互作用
薬物療法, 37 (3) (2017), pp.261-267, 10.1002/phar.1890
スコープで見るGoogle Scholar
フラワーズら、2020年
S.A.フラワーズ、K.M.ウォード、C.T.クラーク
双極性障害における腸内細菌叢と薬物療法管理
Neuropsychobiology, 79 (1) (2020), 43-49頁, 10.1159/000504496
スコープで見るGoogle Scholar
Forslundら、2015年
K. フォルスルンド、F.ヒルデブランド、T.ニールセン、G.ファロニー、E.ル・シャトリエ、砂川聡、O.ペデルセン
ヒト腸内細菌叢における2型糖尿病とメトホルミン治療のシグネチャーの分離
Nature, 528 (7581) (2015), 262-266頁, 10.1038/nature15766
スコープで見るGoogle Scholar
フリードバーグら、2015
D.E.フリードバーグ、N.C.トゥーサン、S.P.チェン、A.J.ラトナー、S.ウィッティア、T.C.ワン、J.A.エイブラムス
プロトンポンプ阻害薬はヒト消化管マイクロバイオームの特定の分類群を変化させる: クロスオーバー試験
e889
Gastroenterology, 149 (4) (2015), pp.883-885, 10.1053/j.gastro.2015.06.043
ScopusGoogle Scholarで見る
藤森, 2015
S. 藤森
プロトンポンプ阻害薬の小腸への影響は?
World J Gastroenterol, 21 (22) (2015), pp.6817-6819, 10.3748/wjg.v21.i22.6817
スコープで見るGoogle Scholar
ゲルハルトとモハジェリ、2018年
S. ゲルハルト、M.H.モハジェリ
パーキンソン病および他の神経変性疾患における大腸細菌組成の変化
ニュートリエンツ, 10 (6) (2018), 10.3390/nu10060708
グーグル・スカラー
ゴマー、2020
E.Z.ゴマー
健康と疾患におけるヒト腸内細菌叢/マイクロバイオーム:総説
Antonie van Leeuwenhoek, 113 (12) (2020), pp.2019-2040, 10.1007/s10482-020-01474-7
Scopusで見るGoogle Scholar
ゴンサレス・アランシビアら、2019年
C. ゴンサレス=アランシビア、J.ウルティア=ピニョネス、J.イラネス=ゴンサレス、J.マルティネス=ピント、R.ソトマイヨール=ザラテ、M.フリオ=ピーパー、J.A.ブラボー
腸内細菌は脳のドーパミンに影響を与えるか?
Psychopharmacology (Berl), 236 (5) (2019), pp.1611-1622, 10.1007/s00213-019-05265-5
Scopusで見るGoogle Scholar
Heら、2022年
D. He, H. Han, X. Fu, A. Liu, Y. Zhan, H. Qiu, X. Wang
メトホルミンは腸内細菌叢を変化させることで治療歴のない2型糖尿病の血糖を低下させる
Can J Diabetes, 46 (2) (2022), 150-156頁, 10.1016/j.jcjd.2021.08.001
PDFで記事を見るScopusで記事を見るGoogle Scholar
北條ら、2018
M. 北條, 浅原利彦, 永原敦子, 武田俊介, 松本和彦, 上山裕之, 渡辺聡子
プロトンポンプ阻害薬使用前後の腸内細菌叢組成
Dig Dis Sci, 63 (11) (2018), pp.2940-2949, 10.1007/s10620-018-5122-4
Scopusで見るGoogle Scholar
ジャクソンら、2016
M.A.ジャクソン、J.K.グッドリッチ、M.E.マキサン、D.E.フリードバーグ、J.A.エイブラムス、A.C.プール、C.J.スティーブス
プロトンポンプ阻害薬は腸内細菌叢の組成を変化させる
Gut, 65 (5) (2016), 749-756頁, 10.1136/gutjnl-2015-310861
スコープで見るGoogle Scholar
金藤ら、2021
H. 金藤秀樹, 木村哲也, 小畑明彦, 下田正樹, 郭一博
長い歴史の中で次々と解明されてきたメトホルミンの多面的作用機序
Int J Mol Sci, 22 (5) (2021), 10.3390/ijms22052596
グーグル奨学生
カーンら、2018
T.J.カーン、Y.M.アーメド、M.A.ザムザミ、A.M.シディキ、I.カーン、O.A.S.バオスマン、M.ヤシール
アトルバスタチン治療は高コレステロール血症患者の腸内細菌叢を調節する
オミックス, 22 (2) (2018), 154-163頁, 10.1089/omi.2017.0130
スコープで見るGoogle Scholar
クヌッセンら、2021年
J.K. Knudsen、C. Bundgaard-Nielsen、S. Hjerrild、R.E. Nielsen、P. Leutscher、S. Sørensen
大うつ病性障害と診断された患者における腸内細菌叢の変動-系統的レビュー
Brain Behav, 11 (7) (2021), Article e02177, 10.1002/brb3.2177
スコープで見るGoogle Scholar
KnueselおよびMohajeri, 2021
T. クヌーセル、M.H.モハジェリ
大うつ病および双極性障害の発症と進行における腸内細菌叢の役割
栄養素, 14 (1) (2021), 10.3390/nu14010037
グーグル奨学生
クーら、2019
S.H. Koo、J. Deng、D.S.W. Ang、J.C. Hsiang、L.S. Lee、S. Aazmi、T.L. Ang
シンガポールにおける多民族グループのヒト腸内細菌叢プロファイルに対するプロトンポンプ阻害薬の影響
Singapore Med J, 60 (10) (2019), pp.512-521, 10.11622/smedj.2018152
Scopusで見るGoogle Scholar
コワルスキーとムラック、2022年
K. コワルスキー、A. ムラク
アルツハイマー病における小腸内細菌の過剰増殖
J Neural Transm (Vienna), 129 (1) (2022), 75-83頁, 10.1007/s00702-021-02440-x
スコープで見るGoogle Scholar
クムメンら、2020年
M. Kummen、O.G. Solberg、C. Storm-Larsen、K. Holm、A. Ragnarsson、M. Trøseid、J.R. Hov
ロスバスタチンはヒト腸内細菌叢の遺伝子組成を変化させる
Sci Rep, 10 (1) (2020), Article 5397, 10.1038/s41598-020-62261-y
Scopusで見るGoogle Scholar
Lagadinouら、2020年
M. Lagadinou、M.O. Onisor、A. Rigas、D.V. Musetescu、D. Gkentzi、S.F. Assimakopoulos、M. Marangos
耐性菌増加時代における非抗生薬の抗菌特性
抗生物質(バーゼル), 9 (3) (2020), 10.3390/antibiotics9030107
Google Scholar
ル・バスタードら、2018
Q. ル・バスタード、G.A.アル=ガリス、M.グレゴワール、G.シャペレ、F.ジャヴァウディン、E.ダイリー、E.モンタシエ
系統的レビュー:抗生物質以外の処方薬によって誘発されるヒト腸内細菌異常症
Aliment Pharmacol Ther, 47 (3) (2018), 332-345頁, 10.1111/apt.14451
スコープで見るGoogle Scholar
リーら、2021年
C.B. Lee, S.U. Chae, S.J. Jo, U.M. Jerng, S.K. Bae
2型糖尿病治療の鍵となる腸内細菌叢とメトホルミンの関係
Int J Mol Sci, 22 (7) (2021), 10.3390/ijms22073566
グーグル奨学生
リーら、2022年
H.-G. リー、M.A.ウィーラー、F.J.キンタナ
神経疾患におけるアストロサイトの機能と治療価値
Nature Reviews Drug Discovery, 21 (5) (2022), pp.339-358, 10.1038/s41573-022-00390-x
スコープで見るGoogle Scholar
リーら、2021年
Y. Lee, A.H. Kim, E. Kim, S. Lee, K.S. Yu, I.J. Jang, J.Y. Cho
腸内細菌叢の変化は、分岐鎖アミノ酸および非必須アミノ酸の代謝の変化を介してメトホルミンの血糖降下作用に影響を及ぼす
Diabetes Res Clin Pract, 178 (2021), Article 108985, 10.1016/j.diabres.2021.108985
PDFで記事を見るScopusで記事を見るGoogle Scholar
李ら、2019
F. Li, P. Wang, Z. Chen, X. Sui, X. Xie, J. Zhang
散発性パーキンソン病の北東漢民族集団における糞便微生物叢の変化
Neurosci Lett, 707 (2019), Article 134297, 10.1016/j.neulet.2019.134297
PDFで記事を見るScopusで記事を見るGoogle Scholar
Liangら、2022年
S. Liang, Z.Y. Sin, J. Yu, S. Zhao, Z. Xi, R. Bruzzone, H.M. Tun
うつ病に関連する腸内細菌のマルチコホート解析により、集団間の細菌バイオマーカーに関する知見が得られた。
Cell Mol Life Sci, 80 (1) (2022), Article 9, 10.1007/s00018-022-04650-2
Scopusで見るGoogle Scholar
リャンら、2018
S. リャン、X.ウー、X.フー、T.ワン、F.ジン
微生物叢-腸-脳軸からうつ病を認識する
Int J Mol Sci, 19 (6) (2018), 10.3390/ijms19061592
グーグル・スカラー
リビー、2020
P. リビー
スタチン製剤が健康な腸内細菌を増やす可能性
ネイチャー誌581巻(2020年)263-264頁、10.1038/d41586-020-01281-0号所収。
スコープで見るGoogle Scholar
リム, 2020
G.B.リム
スタチン服用中の肥満患者における腸内細菌叢プロファイルの改善
ネイチャー・レビュー心臓病学、17 (7) (2020)
385-385. https://doi.org/10.1038/s41569-020-0396-6
Google Scholar
林ら、2021年
Y.T. Lin, T.Y. Lin, S.C. Hung, P.Y. Liu, P.H. Wu, Y.S. Chuang, C.Y. Wu
抗酸菌薬投与は血液透析患者の腸内細菌叢組成に変化をもたらす
微生物, 9 (2) (2021), 10.3390/microorganisms9020286
グーグル奨学生
劉ら、2022
W. 劉、羅、周、孫
腸内細菌叢と抗糖尿病薬: 2型糖尿病における個別化治療の展望
Front Cell Infect Microbiol, 12 (2022), Article 853771, 10.3389/fcimb.2022.853771
スコープで見るGoogle Scholar
マセドら、2017
D. マセド、A.J.M.C.フィリョ、C.N.ソアレス・デ・ソウザ、J.ケベド、T.バリチェロ、H.V.N.ジュニオール、D.フレイタス・デ・ルセナ
抗うつ薬、抗菌薬、あるいはその両方?うつ病における腸内細菌叢の異常と抗うつ薬の抗菌作用が抗うつ薬の有効性に及ぼす影響の可能性
Journal of Affective Disorders, 208 (2017), 22-32頁, 10.1016/j.jad.2016.09.012
PDFで記事を見るScopusで記事を見るGoogle Scholar
マッケら、2020年
L. マッケ、C.シュルツ、L.コレツコ、P.マルファータイナー
系統的レビュー:消化管のマイクロバイオームに対するプロトンポンプ阻害薬の影響-次世代シーケンス研究からのエビデンス
Aliment Pharmacol Ther, 51 (5) (2020), 505-526頁, 10.1111/apt.15604
スコープで見るGoogle Scholar
Mahmoudら、2019
S. マフムード、M.ガラゴズルー、C.シマール、D.グリ
アストロサイトはグルタミン酸の取り込みと放出のバランスを制御することで、中枢神経系におけるグルタミン酸の恒常性を維持する
Cells, 8 (2) (2019), 10.3390/cells8020184
グーグル・スカラー
マニアールら、2017
K. マニアール、A.モイディーン、R.バタチャリヤ、D.バネルジー
メトホルミンは糖尿病患者において腸内細菌叢の調節を介して抗肥満効果を発揮する: 仮説
Med Hypotheses, 104 (2017), 117-120頁, 10.1016/j.mehy.2017.06.001
PDFで記事を見るScopusで記事を見るGoogle Scholar
マセダとリチョッティ、2020年
D. マセダ、E. リッチョッティ
非ステロイド性抗炎症薬と腸内細菌叢の相互作用
Front Pharmacol, 11 (2020), Article 1153, 10.3389/fphar.2020.01153
スコープで見るGoogle Scholar
マクギネスら、2022年
A.J. McGuinness、J.A. Davis、S.L. Dawson、A. Loughman、F. Collier、M. O'Hely、F.N. Jacka
大うつ病性障害、双極性障害、統合失調症の観察研究における腸内細菌叢組成の系統的レビュー
Mol Psychiatry, 27 (4) (2022), 1920-1935頁, 10.1038/s41380-022-01456-3
スコープで見るGoogle Scholar
医学, 2022
医学、J.H.(2022)。どのようにスタチン薬は心臓を保護する。で。
Google Scholar
ミナリヤンら、2017
A. Minalyan, L. Gabrielyan, D. Scott, J. Jacobs, J.R. Pisegna
胃と腸のマイクロバイオーム: プロトンポンプ阻害薬の役割
Curr Gastroenterol Rep, 19 (8) (2017), Article 42, 10.1007/s11894-017-0577-6
スコープで見るGoogle Scholar
モライスら、2021年
L.H.モライス、H.L.tシュライバー、S.K.マズマニアン
行動と脳疾患における腸内細菌叢-脳軸
Nat Rev Microbiol, 19 (4) (2021), pp.241-255, 10.1038/s41579-020-00460-0
スコープで見るGoogle Scholar
ミューラーら、2021年
N.T. Mueller、M.K. Differding、M. Zhang、N.M. Maruthur、S.P. Juraschek、E.R. Miller 3rd, H.C. Yeh
メトホルミンは腸内細菌叢の組成と機能および循環短鎖脂肪酸に影響する:無作為化試験
糖尿病ケア, 44 (7) (2021), 1462-1471頁, 10.2337/dc20-2257
スコープで見るGoogle Scholar
内藤ら、2018
Y. 内藤, 柏木健一郎, 高木崇, 安藤晃, 井上玲子
プロトンポンプインヒビター使用による腸内細菌異常症
消化, 97 (2) (2018), pp.195-204, 10.1159/000481813
スコープで見るGoogle Scholar
ヌザムら、2020年
N.D. Nuzum、A. Loughman、E.A. Szymlek-Gay、A. Hendy、W.P. Teo、H. Macpherson
健康な高齢者とパーキンソン病患者の腸内細菌叢の違い: 系統的レビュー
Neurosci Biobehav Rev, 112 (2020), 227-241頁, 10.1016/j.neubiorev.2020.02.003
PDFで記事を見るScopusで記事を見るGoogle Scholarで記事を見る
OECD iLibrary, 2021
OECD iLibrary, O.H. S. (2021). 医薬品消費。〈https://www.oecd-ilibrary.org/sites/5689c05c-en/index.html?itemId=/content/component/5689c05c-en〉.
Google Scholar
オンライン、2022年
オンライン、D. (2022). 医薬品統計。In.
Google Scholar
組織、2021年
組織、W.H.(2021)。必須医薬品モデルリスト2021(22リスト)。In.
Google Scholar
大谷ら、2017
K. 大谷和彦, 谷川俊彦, 渡辺哲也, 島田聡, 灘谷康弘, 永見祐子, 荒川哲也
微生物叢は非ステロイド性抗炎症薬誘発小腸障害に重要な役割を果たす
消化, 95 (1) (2017), pp.22-28, 10.1159/000452356
スコープで見るGoogle Scholar
ペッレグリーニら、2018
C. ペッレグリーニ、L.アントニオーリ、R.コルッチ、C.ブランディッツィ、M.フォルナイ
腸内細菌叢、腸粘膜バリア、腸管神経免疫系の相互作用:神経変性疾患への共通経路?
Acta Neuropathol, 136 (3) (2018), pp.345-361, 10.1007/s00401-018-1856-5
Scopusで見るGoogle Scholar
Rajputら、2021年
C. ラジプット、A.サルカール、N.サチャン、N.ラワット、M.P.シン
腸内細菌異常症はパーキンソン病の震源地か?
Neurochem Res, 46 (3) (2021), pp.425-438, 10.1007/s11064-020-03187-9
スコープで見るGoogle Scholar
ライチェルとクナウフ、2021年
M. ライチェル、F. クナウフ
スタチン、肥満、マイクロバイオーム:スタチン治療の多面的効果の潜在的メカニズム
Kidney International, 99 (3) (2021), pp.531-533, 10.1016/j.kint.2020.07.038
PDFで記事を見るScopusで記事を見るGoogle Scholar
ロジャースら、2013
M.A.ロジャース、M.T.グリーン、V.B.ヤング、S.セイント、K.M.ランガ、J.Y.カオ、D.M.アロノフ
うつ病、抗うつ薬、クロストリジウム・ディフィシル感染症のリスク
BMC Med, 11 (2013), Article 121, 10.1186/1741-7015-11-121
Scopusで見るGoogle Scholar
ロジャースとアロノフ、2016
M.A.M.ロジャース、D.M.アロノフ
非ステロイド性抗炎症薬が腸内細菌叢に及ぼす影響
178.e171-178.e179
Clin Microbiol Infect, 22 (2) (2016), 10.1016/j.cmi.2015.10.003
グーグル・スカラー
ライマンら、2023年
S. ライマン、A.A.ヴァフチン、S.P.リチャードソン、H.C.リン
前駆期および症候性レビー小体病における微生物-腸-脳の機能障害
J Neurol, 270 (2) (2023), 746-758頁, 10.1007/s00415-022-11461-9
スコープで見るGoogle Scholar
Shenら、2021
Y. Shen、X. Yang、G. Li、J. Gao、Y. Liang
SSRI治療下のMDD患者における腸内細菌叢の変化
Sci Rep, 11 (1) (2021), Article 14918, 10.1038/s41598-021-94481-1
スコープで見るGoogle Scholar
シドゥおよびファン・デル・ポールテン、2017年
M. シドゥ、D.ファン・デル・ポールテン
腸内マイクロバイオーム
Aust Fam Physician, 46 (4) (2017), pp.
スコープで見るGoogle Scholar
シングルケア.
シングルケア.(2022). 処方薬統計2022年。In.
グーグル・スカラー
スコニエチュナ・ジデツカら、2018年
K. Skonieczna-Żydecka, E. Grochans, D. Maciejewska, M. Szkup, D. Schneider-Matyka, A. Jurczak, E. Stachowska
ポーランドのうつ病女性における糞便中短鎖脂肪酸プロファイルの変化
Nutrients, 10 (12) (2018), 10.3390/nu10121939
グーグル・スカラー
ソフトウェア, 2023
ソフトウェア、B.https://www.biorender.com/〉。
Google Scholar
ステートメント、2020年
ステートメント、P.系統的レビューとメタアナリシスのための望ましい報告項目。〈http://www.prisma-statement.org/PRISMAStatement/FlowDiagram〉。
グーグル・スカラー
Statista.com, 2022
Statista.(2022). 米国における2004年から2020年までのメトホルミン処方数。In.
グーグル・スカラー
ストランドヴィッツ、2018
P. ストランドヴィッツ
腸内細菌叢による神経伝達物質調節
Brain Res, 1693 (Pt B) (2018), pp.128-133, 10.1016/j.brainres.2018.03.015
PDFで記事を見るScopusで記事を見るGoogle Scholar
サンら、2019
L. スン、チャン、カオ、ワン、チャオ、ワン、ニー、Y.
フルオキセチンは、マウスの慢性的な予期せぬ軽度のストレスによって誘発されるうつ病モデルにおけるディスバイオーシスを改善する
Int J Med Sci, 16 (9) (2019), pp.1260-1270, 10.7150/ijms.37322
スコープで見るGoogle Scholar
シリオンら、2023
F. Thirion, F. Sellebjerg, Y. Fan, L. Lyu, T.H. Hansen, N. Pons, O. Pedersen
多発性硬化症における腸内細菌叢は疾患活動性によって異なる
Genome Med, 15 (1) (2023), Article 1, 10.1186/s13073-022-01148-1
スコープで見るGoogle Scholar
トンら、2018
X. トン、J.シュー、F.リアン、X.ユー、Y.ザオ、L.シュー、L.ザオ
メトホルミンと漢方処方による高脂血症を伴うヒト2型糖尿病の改善における腸内細菌叢の構造変化:多施設共同無作為化オープンラベル臨床試験
mBio, 9 (3) (2018), 10.1128/mBio.02392-17
グーグル・スカラー
トランとモハジェリ、2021年
S.M.トラン、M.H.モハジェリ
脳の発達、老化、疾患における腸内細菌代謝産物の役割
栄養素, 13 (3) (2021), 10.3390/nu13030732
グーグル奨学生
トルバら、2021
R. Troubat、P. Barone、S. Leman、T. Desmidt、A. Cressant、B. Atanasova、V. Camus
神経炎症とうつ病: 総説
Eur J Neurosci, 53 (1) (2021), 151-171頁, 10.1111/ejn.14720
スコープで見るGoogle Scholar
タイラー・パターソンとグランディ, 2020
T. タイラー・パターソン、R. グランディ
腸内細菌叢と神経疾患・損傷
Adv Exp Med Biol, 1238 (2020), 73-91頁, 10.1007/978-981-15-2385-4_6
Scopusで見るGoogle Scholar
宇津井・宇津井、2017年
E. ウツェリ、P. ウサイ
腸管透過性と非アルコール性脂肪肝疾患に対する非ステロイド性抗炎症薬の役割
World J Gastroenterol, 23 (22) (2017), pp.3954-3963, 10.3748/wjg.v23.i22.3954
スコープで見るGoogle Scholar
バジェス・コロマーら、2019年
M. バジェス・コロマー、G.ファロニー、Y.ダージ、E.F.ティグチェラー、J.ワン、R.Y.ティト、J.レーズ
QOLとうつ病におけるヒト腸内細菌叢の神経活性の可能性
Nat Microbiol, 4 (4) (2019), 623-632頁, 10.1038/s41564-018-0337-x
Scopusで見るGoogle Scholar
ヴァリアヌーら、2019年
N.G.ヴァリアヌー、T.ストラティグー、S.ツァガラキス
メトホルミンと腸内細菌叢:それらの相互作用と糖尿病への影響
ホルモン(アテネ), 18 (2) (2019), pp.141-144, 10.1007/s42000-019-00093-w
Scopusで見るGoogle Scholar
ヴィア・メディチ、2023年
via medici, T. (2023). Orale Antidiabetika und GPL-1 Analoga. 〈https://viamedici.thieme.de/lernmodul/8659126/4915659/orale+antidiabetika+und+glp-1-analoga#_EBDA5E30_5F9F_49EF_BBCA_A0D7A3D459BC〉
Google Scholar
ヴィエイラ・シルヴァら、2020年
S. ヴィエイラ-シルヴァ、G.ファロニー、E.ベルダ、T.ニールセン、J.アロン-ウィスネフスキー、R.チャカロン、J.レーズ
スタチン治療は腸内細菌叢異常症の有病率の低下と関連する
Nature, 581 (7808) (2020), 310-315頁, 10.1038/s41586-020-2269-x
スコープで見るGoogle Scholar
Wangら、2021年
X. Wang, H. Huang, Y. Zhu, S. Li, P. Zhang, J. Jiang, J. Lai
メトホルミンは腸脳軸に作用し、抗精神病薬による代謝機能障害を改善する
Biosci Trends, 15 (5) (2021), pp.321-329, 10.5582/bst.2021.01317
スコープで見るGoogle Scholar
王ら、2021年
X. Wang, Q. Tang, H. Hou, W. Zhang, M. Li, D. Chen, H. Cao
非ステロイド性抗炎症薬腸症における腸内細菌叢:内部からの新たな知見
Front Cell Infect Microbiol, 11 (2021), Article 679396, 10.3389/fcimb.2021.679396
スコープで見るGoogle Scholar
ワウタースら、2021年
L. Wauters、R.Y.Tito、M.Ceulemans、M.Lambaerts、A.Accarie、L.Rymenans、J.Raes
機能性ディスペプシアにおけるプロトンポンプ阻害薬の有効性と十二指腸ディスバイオーシスとの関係
Int J Mol Sci, 22 (24) (2021), 10.3390/ijms222413609
グーグル奨学生
ワイスとヘネット, 2017
G.A.ワイス、T.ヘネット
腸内細菌異常症のメカニズムと結果
細胞分子生命科学, 74 (16) (2017), pp.2959-2977, 10.1007/s00018-017-2509-x
Scopusで見るGoogle Scholar
ウィンターら、2018
G.ウィンター、R.A.ハート、R.P.G.チャールズワース、C.F.シャープリー
腸内マイクロバイオームとうつ病:わかっていることと知るべきこと
Rev Neurosci, 29 (6) (2018), 629-643頁, 10.1515/revneuro-2017-0072
Scopusで見るGoogle Scholar
ヴィットゥング-シュターフシェーデ、2022
P. ヴィットゥング=シュターフシェーデ
腸の力: 消化管におけるアミロイド生成タンパク質によるヒトアミロイド形成の調節
Curr Opin Struct Biol, 72 (2022), 33-38頁, 10.1016/j.sbi.2021.07.009
PDFで記事を見るScopusで記事を見るGoogle Scholar
ウーら、2017
H. Wu, E. Esteve, V. Tremaroli, M.T. Khan, R. Caesar, L. Mannerås-Holm, F. Bäckhed
メトホルミンは未治療の2型糖尿病患者の腸内細菌叢を変化させ、薬剤の治療効果に寄与する
Nature Medicine, 23 (7) (2017), 850-858頁, 10.1038/nm.4345
スコープで見るGoogle Scholar
Wuら、2020
H. Wu, V. Tremaroli, C. Schmidt, A. Lundqvist, L.M. Olsson, M. Krämer, F. Bäckhed
糖尿病前症および糖尿病における腸内細菌叢: 集団ベースの横断研究
e373
細胞代謝, 32 (3) (2020), 379-390頁, 10.1016/j.cmet.2020.06.011
スコープで見るGoogle Scholar
Wuら、2022年
Y. Wu, Z. Hang, T. Lei, H. Du
腸内細菌叢は内因性ホルモンを制御することでアルツハイマー病に影響を及ぼす
Neurochem Res, 47 (12) (2022), 3565-3582頁, 10.1007/s11064-022-03784-w
スコープで見るGoogle Scholar
Yaoら、2023
H. Yao, D. Zhang, H. Yu, H. Shen, H. Liu, F. Meng, X. Wang
うつ病の病態における微生物叢-腸-脳軸: ナラティブレビュー
Physiol Behav, 260 (2023), Article 114056, 10.1016/j.physbeh.2022.114056
PDFで記事を見るScopusで記事を見るGoogle Scholar
Zhangら、2022年
K. Zhang、K.C.Paul、J.P.Jacobs、H.L.Chou、A.Duarte Folle、I.Del Rosario、B.Ritz
カリフォルニア農村部におけるパーキンソン病と腸内マイクロバイオーム
J Parkinsons Dis, 12 (8) (2022), 2441-2452頁, 10.3233/jpd-223500
スコープで見るGoogle Scholar
張と胡、2020年
Q. 張、胡
肥満および2型糖尿病における腸内細菌叢に対するメトホルミンの効果
Diabetes Metab Syndr Obes, 13 (2020), pp.5003-5014, 10.2147/dmso.s286430
スコープで見るGoogle Scholar
Zhangら、2021
W. Zhang、W. Qu、H. Wang、H. Yan
抗うつ薬フルオキセチンとアミトリプチリンは、予測不可能な軽度ストレスに慢性的に曝露したラットにおいて、腸内細菌叢と腸内マイクロバイオーム機能の変化を誘導する
Transl Psychiatry, 11 (1) (2021), Article 131, 10.1038/s41398-021-01254-5
Google Scholar
Zhao et al.
H. Zhao, K. Jin, C. Jiang, F. Pan, J. Wu, H. Luan, M. Huang
大うつ病性障害における腸内細菌叢のマルチオミクス研究の試験的探索
Transl Psychiatry, 12 (1) (2022), Article 8, 10.1038/s41398-021-01769-x
スコープで見るGoogle Scholar
周ら、2022年
R. Zhou、S. Qian、W.C.S. Cho、J. Zhou、C. Jin、Y. Zhong、H. Zhang
加齢に伴う認知機能低下における微生物叢とミクログリアのつながり
Aging Cell, 21 (5) (2022), Article e13599, 10.1111/acel.13599
スコープで見るGoogle Scholar
ザドリら、2023年
Z.S. Zádori、K. Király、M. Al-Khrasani、K. Gyires
非ステロイド性抗炎症薬、オピオイドと腸内細菌叢の相互作用-炎症と疼痛管理における将来の展望
Pharmacology & Therapeutics, 241 (2023), Article 108327, 10.1016/j.pharmthera.2022.108327
PDFで記事を見るScopusで記事を見るGoogle Scholar
引用者: (0)
© 2024 エルゼビア社発行
エルゼビアのロゴとワードマーク
サイエンスダイレクトについて
リモートアクセス
ショッピングカート
広告掲載
お問い合わせとサポート
利用規約
プライバシーポリシー
当サイトではクッキーを使用しています。クッキー設定
このサイトのすべてのコンテンツ: Copyright © 2024 Elsevier B.V., its licensors, and contributors. テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビアに帰属します。すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条項が適用されます。
RELXグループホームページ
フィードバック
この記事が気に入ったらサポートをしてみませんか?