乳酸菌発酵生薬の免疫調節特性に関する新たな知見

Front. Microbiol.、2022年11月28日
Sec. Food Microbiology
https://doi.org/10.3389/fmicb.2022.1073922
乳酸菌発酵生薬の免疫調節特性に関する新たな知見
朱宏儒、郭麗東、兪丹、杜小維*(英語名:Xiaowei Du
黒龍江中医薬大学薬学院(中国・ハルビン市


COVID-19の大流行により、感染症に対する身体の防御システムである免疫システムに注目が集まっている。伝統的な漢方薬の免疫調節能力は臨床試験研究によって確認されており、潜在的な標的の数が多く、毒性が低いため、処方薬よりも明らかに有利である。生薬の活性化合物には、主に多糖類、サポニン、フラボノイド、フェノール類などがあり、乳酸菌(LAB)発酵後に修飾して新しい活性化合物を生成することができる。プロバイオティクスの主要供給源であるLABは、エクソポリサッカライド、短鎖脂肪酸、バクテリオシンなどの免疫調節代謝産物をさらに産生することが可能である。さらに、漢方薬に含まれるいくつかの化合物は、LABベースの免疫活性代謝物の増殖と生産を促進することができる。このように、LABを介した生薬の発酵は、ヒトの免疫反応を調節するための新たな戦略となってきている。今回の総説では、LAB発酵生薬の免疫調節特性および活性化合物、LABと生薬の相互作用、発酵中に起こる免疫調節成分の変化について論じている。また、自然免疫系または適応免疫系の活性化、腸管免疫恒常性の維持など、LAB発酵生薬が免疫反応を調節するメカニズムについても考察している。
はじめに
ヒトの免疫系は、健康な組織を攻撃したり、有益な生物を危険にさらさないようにしながら、外来病原体から体を守るために徐々に進化してきた。免疫系は、リンパ系器官、細胞、液性因子、サイトカインの相互作用ネットワークから成り、免疫監視、防御、安定化という3つの主要機能を有している(Chaplin, 2010)。免疫系が正常に機能していれば、侵入してきたウイルス、老化細胞、腫瘍細胞などを確実に識別し、除去することができる。ここ数年の研究により、病気の発生や進展は、免疫機能と密接に関係していることが明らかになっています。免疫活性の低下は、重症感染症、がん、免疫不全症に関連し、活性の上昇は、アレルギー性疾患、炎症性疾患、自己免疫疾患につながります(Levy et al.、2017)。COVID-19の病態は、典型的には、感染に対する免疫反応によって引き起こされる急性の進行性肺微小循環障害を含んでいます。研究によると、罹患率および死亡率は、高齢者および免疫不全者において高いことが示されています(Forouzani-Haghighi et al, 2022)。このように、免疫系はCOVID-19の病因と予後において重要な役割を担っていることが明らかになっています。
漢方薬は、様々な病状の予防や治療に有効であることから、常用されています。生薬には、多糖類、トリテルペン、フラボノイド、アルカロイド、揮発性油、フェノールなど多くの活性化合物があるが、特定の活性化合物の構造上、人体が効果的に吸収することは難しく、従来の方法で精製することも同様に困難であった。科学者たちは、生薬中の活性物質の含有量とバイオアベイラビリティを向上させるために、さまざまな物理的、化学的、生物変換的な方法を用いてきた。近年、微生物変換技術は、その強い特異性、限られた副産物、穏やかな反応条件、環境に優しいプロセスから、天然医薬品の変換に広く使用されています。微生物変換される微生物としては、真菌と細菌が最も一般的である。例えば、Dioscorea nipponicaから分離された内生菌であるFusarium sp. C39は、Dioscoreaサポニンをジオスゲニンに効果的に変換できる。このプロセスには、解糖代謝、閉ループ反応、脱水素化、およびカルボニル化が含まれると考えられる(Huang et al.、2022)。さらに、ハトムギ(Coix lacryma-jobi)の枯草菌媒介発酵は、テトラメチルピラジン、γ-アミノ酪酸、トリテルペン、フェノール、フラボノイドおよび、コイクセノリドのレベルを著しく増加することができる(Wen et al.、2020年)。β-グルコシダーゼ産生菌は、希少なサポニン、イソフラボンなどの生体内変換研究によく選ばれる(Geraldi et al, 2020; Lodha et al., 2021)。乳酸菌(LAB)は、数ある代替発酵菌株の中でも、その安全性と効率性から際立っている。
乳酸菌は、食品発酵および生物医学産業で広く使用されている一般に安全と認められた(GRAS)微生物である。乳酸菌発酵は、酵素の作用により望ましくない基質を分解し、新しい活性物質に変換することができるため、天然医薬品の生物活性を向上させることができる。例えば、Lactiplantibacillus plantarumおよびLeuconostoc mesenteroidesを用いた乾燥ロンガン果肉発酵は、フリーおよび総フェノール含有量を増加させることによってその抗酸化活性を高めることができる(Khanら、2018)。さらに、Lp.plantarumを介した発酵は、α-グリコシダーゼの阻害活性を高め、DPPHのラジカル消去および抗糖化を抑制することによって、伝統的なハーブ混合物であるDanggui Buxue Tang (DBT) の2型糖尿病に対する治療効果を向上させる。発酵DBTは、非発酵DBTとは異なるフラボノイド化合物を生成することが示されている(Guo et al.、2020)。また、一部の天然化合物は、微生物の代謝経路に影響を与え、活性二次代謝産物の量を増加させる。
最近の研究では、プロバイオティクス細菌とその発酵生成物の人間の健康に対する有益な作用に焦点が当てられている。LABは、ヒト免疫細胞と相互作用し、様々な炎症性疾患に対する自然免疫反応および適応免疫反応に関与する特定の経路を調節することが示されている(Zhao et al.、2019)。特に、発酵システムにおけるプレおよびプロバイオティクスは、ヒトの腸の微生物および活性に好影響を与え、免疫系をさらにサポートします(Peters V. et al., 2019)。これらの理由から、生薬のLAB発酵は、生薬の免疫調節活性を高めるために有用である。本研究では、LAB発酵が生薬の有効成分に及ぼす影響と、生薬が免疫系を活性化するメカニズムについて概説する。
乳酸菌発酵生薬の免疫調節効果
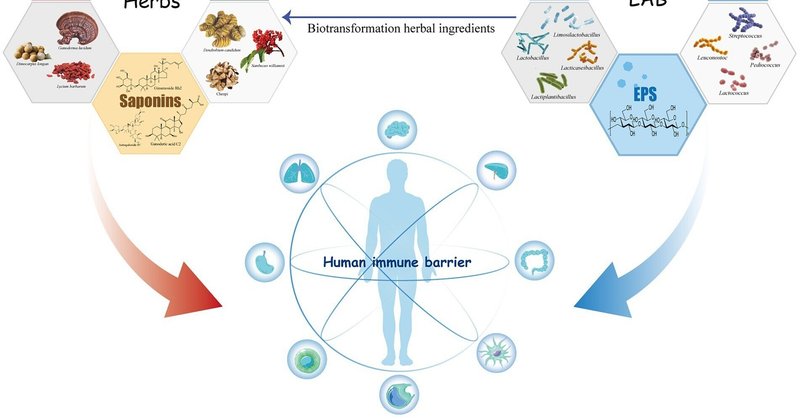
乳酸菌は、発酵中の生薬成分の免疫調節活性に好影響を与える。乳酸菌の代謝活性は、生薬中の有効成分の含有量や構造を変化させる。天然物も同様に、乳酸菌の増殖状態や代謝過程に影響を与える(図1)。
図1
www.frontiersin.org
図1. 乳酸菌と生薬の免疫調節活性成分とその相互作用。
生薬活性成分の免疫調節作用
生薬には古くから、腫瘍、感染症、自己免疫疾患など、人間の多くの疾患に有効な天然のドラッグライブラリーが含まれていると考えられてきた。ハーブに含まれる多くの成分は、免疫反応を活性化することによって、健康に役立つことが示されている。例えば、霊芝、リュウキュウアサガオ、高麗人参、レンゲ、デンドロビウムなどのハーブに含まれる多糖類、サポニン、ポリフェノールは、強い免疫調節活性を有しています。
植物・菌類多糖類
Astragalus membranaceus, Longan, and L. barbarum (Yin et al., 2019) の植物性多糖類、および G. lucidum と Poria cocos (Yin et al., 2021) の真菌性多糖類などの多糖類は、生薬における共通の巨大分子免疫調節物質である。いくつかの研究により、植物多糖類は免疫系に対してマルチチャンネルおよびマルチレベルの調節作用を有することが示されている。それらはT細胞、B細胞、マクロファージ、ナチュラルキラー細胞などのいくつかの免疫細胞を活性化し、補体を活性化し、サイトカインの産生を促進し、それによって免疫系の多くの構成要素を制御することができる。例えば、植物多糖類は、細胞表面の特定の受容体を認識して結合することにより、マクロファージを活性化し、それによって免疫応答を開始することができる(Yin et al.、2019)。G. lucidumまたは特定の食用菌からの菌類β-グルカンは、免疫細胞を活性化することにより、間接的にがん細胞または腫瘍に抵抗することができます。また、βグルカンは、炎症性サイトカインであるIL-6やTNF-αを減少させ、抗酸化物質の生産を増加させることでアレルギーを緩和する(Murphyら、2020)、パターン認識受容体が引き起こす作用機構と類似している。
サポニン
サポニンは、薬草に含まれる二次代謝産物群の一つで、様々なメカニズムで免疫調節作用を示す。サポニン化合物は、自己免疫疾患、アレルギー性疾患、癌の治療に用いられている。例えば、アストラガロシドIVは、単球のM1マクロファージへの分化を誘導し、抗腫瘍免疫反応を開始させることができる(Min et al, 2022)。グリチルレチン酸は、Th2サイトカインであるIL-5およびIL-13の産生を抑制することにより、喘息マウスモデルにおけるアレルギー反応を緩和することができる(Kim et al.、2017年)。ジンセノサイドRh2は、腫瘍におけるTリンパ球の浸潤および細胞毒性を増強し、担癌マウスの細胞性免疫機能を改善することができる(Wang et al.、2017)。ジンセノサイド化合物K(CK)は、膠原病性関節炎マウスのTリンパ球の異常活性化を抑制し、メモリーT細胞を抑制して、免疫系が心臓移植を拒絶するのを防ぐことができる(Wangら、2009年;Liuら、2014年)。
ポリフェノール
ポリフェノールは、3員環のフラバン構造を持つ植物由来の有機化合物です。これらの物質は、フリーラジカルの消去、酸化ストレスの低下、炎症の軽減など、健康増進に役立つ性質が知られています(Abbas et al.、2017)。抗酸化剤および抗炎症剤として作用するフェノール化合物は、酸化ストレスを緩和し、腸の炎症反応の動的平衡を保ち、免疫学的耐性を促進するのに役立ちます(Williamson, 2017)。フラボノイドとフェノール酸は、ポリフェノールの主要なファミリーメンバーです。フラボノイドは、自己免疫疾患や癌を含むいくつかの病気に対する免疫調節効果の可能性があります(Ginwala et al.、2019)。また、免疫細胞の主要な酵素機能を調節し、免疫調節活性を媒介することが分かっています(D'Arcy, 2022)。レスベラトロールなどの他の天然フェノール化合物も優れた免疫賦活活性を示し、免疫細胞の浸潤を増加させ、表現型を調節し、関連サイトカインの分泌を促進します(Alesciら、2022年)。
生理活性アミノ酸
ガンマアミノ酪酸(GABA)は、天然のリラックスアミノ酸の一種で、不安、うつ、高血圧を抑制し、ホルモン分泌を調整する働きがあります。GABAは、マメ科の植物や高麗人参などの種子、根茎、組織液、生薬などに含まれています。食事性GABAは非特異的免疫を改善し、青年期のリポポリサッカライドによって誘発される免疫亢進を抑制します(Zhangら、2022年)。先行研究では、低生理濃度GABAがT細胞特異的GABAチャネルを活性化することでT細胞の増殖を抑制し、免疫反応を抑制することが示されており、この役割は自己免疫疾患の治療において有望である(Bjurstöm et al.2008; Prud'home et al.2015 )。また、別の研究では、GABAの経口投与がストレス条件下での免疫反応を高めることがわかりました(Abdou et al.、2006)。このように、GABAを適切に補給することは、人間の健康にとって有益であると考えられます。
乳酸菌の免疫調節作用と活性物質
乳酸菌は、免疫調節作用をはじめ、ヒトの健康に対していくつかの有益な作用を有しています。これらの機能は、動物モデルやヒト臨床試験で確認されており、細胞外多糖類や短鎖脂肪酸などの乳酸菌代謝物の活性と密接な関係があることが示されている。
細胞外多糖類
プロバイオティクス細菌は、様々な構造・組成・機能を持つ多糖類を外界に分泌している。これらはエキソポリサッカライド(EPS)と呼ばれ、その多くは抗腫瘍、免疫調節、血糖コントロール、抗酸化など幅広い効果を有している(Nwodoら、2012)。EPSの免疫調節機能は、これらの細菌のエキソ多糖の化学構造と構成に強く相関している(Rajoka et al.) 具体的には、乳酸菌のEPSは、水酸基、リン酸基、カルボニル基などを含み、これらの基が生物活性を促進する(Rajoka et al.、2020)。Lacticaseibacillus rhamnosus KL37からのEPSに応答して、マウス腹膜マクロファージは、TNF-α、IL-6、およびIL-12のような炎症性サイトカイン、ならびにIL-10のような抗炎症性サイトカインを生成し、これらの化合物が著しい免疫活性を引き起こすことを示す(Ciszek-Lendaら, 2011)。別の研究では、Lp. Plantarum JLAU10から酸性細胞外多糖を単離・精製し、この多糖がLp. Plantarum JLAU103、EPS103と名付けられ、強い二重免疫調節活性を有する(Wang et al.、2020)。EPS103は、RAW264.7マクロファージを活性化すると同時に、IL-6、TNF-α、プロスタグランジンE2(PGE2)、NO、シクロオキシゲナーゼ2(COX-2)、誘導性NO合成酵素(iNOS)などのいくつかの炎症因子の過剰放出とmRNA発現を低減することが判明した(Wangら,2020年)。これらの結果は、LAB特異的EPSが免疫調節物質として作用する可能性を示唆している。
短鎖脂肪酸
大腸の嫌気性細菌は、難消化性の食物繊維を酢酸、プロピオン酸、酪酸などの短鎖脂肪酸(SCFAs)に発酵させます。これらは、エネルギー源として機能し、様々なシグナル伝達プロセスに関与しています(Russell et al.、2013)。最近の研究では、微生物叢由来のSCFAは、Gタンパク質受容体41(GPR41)およびヒストン脱アセチル化酵素(HDAC)を阻害することにより、CD4+ T細胞および自然リンパ系細胞(ILC)からのIL-22産生の誘導に重要な役割を果たし、腸の恒常性を維持することが分かった(Yang et al.、2020年)。別の研究では、乳酸とSCFAの両方が、TLRシグナル伝達を用量依存的に阻害することによって、腸管上皮細胞および骨髄系細胞を含むセンチネル細胞における炎症性応答をダウンレギュレートし、それによって胃腸の恒常性を回復できることが分かった(Iraporda et al.、2015年)。SCFAであるペンタノエートは、IL-10の産生を促進し、Th17細胞を抑制することにより、マウスの腸および脳におけるT細胞媒介性免疫病理を低減することが示された。したがって、ペンタノエートは、炎症性疾患および自己免疫疾患の治療において可能性がある(Luuら、2019)。
バクテリオシン(Bacteriocins
LABは増殖する際に、バクテリオシンと呼ばれる抗菌タンパク質またはポリペプチドを作り、食品保存料と食中毒の治療薬の両方として使用されてきた(Zacharof and Lovitt, 2012)。バクテリオシンは、治療用の抗生物質や免疫調節剤として使用される可能性があり、注目を集めている(Huangら、2021)。プロバイオティクスが産生するバクテリオシンは、免疫系にも好影響を与えることができる。病原体に直接拮抗することで宿主の炎症反応を抑えるほか、一部のバクテリオシンは免疫細胞を活性化し、免疫力を高めることができます(Peters V. et al., 2019)。Lactococcus lactisが産生するナイシンは、マクロファージ誘導IL-12産生を増強し(Moein et al., 2018)、炎症因子のレベルを双方向的に変更し、免疫バランスを維持することが示されている(Małaczewska et al., 2019)。S結合型糖ペプチドであるスブランシンは、マクロファージの機能を高め、CD4+およびCD8+細胞の増殖を促進することもわかっています(Wangら、2018;Wang J.ら、2019)。この糖ペプチドは、核因子-κB(NF-κB)の活性化を抑制して腸の炎症を和らげることができます(Hu et al.、2018)。Gタンパク質共役型ヒドロキシカルボン酸受容体(HCAR)は、代謝や食事条件が変化すると、免疫機能やエネルギーバランスを調節します(Offermanns, 2017)。LABが生産する抗菌性代謝物であるD-3-フェニル乳酸(D-PLA)は、ヒトおよび類人猿特異的なHCA3受容体を特異的に認識し、免疫細胞にシグナルを伝達して免疫応答を誘導する(Peters A. et al.、2019)。しかし、D-PLAによるHCA3活性化が単球の貪食やマクロファージへの分化に影響を与えるかどうかは、さらなる研究が必要である。
乳酸菌と生薬の相互作用。免疫調節の促進効果
乳酸菌と生薬成分は、発酵中に双方向に相互作用する。この過程で、生薬中の様々な免疫反応性化合物の合成を誘導したり、天然分子を修飾して免疫調節効果を高めたりして、天然物の生物活性を向上させる(表1)。同時に、ハーブに含まれる成分の中には、LABの増殖や有益な代謝産物の生産を促進するものもある。LAB発酵と生薬の関係、およびそれらが宿主の免疫系に与える影響を図1に示す。
表1
www.frontiersin.org
表1. 乳酸菌発酵させた生薬の免疫調節効果。
乳酸菌発酵による生薬への影響
植物・菌類の多糖類に対する発酵の効果
発酵は、植物や菌類の多糖体含量を増加させ、植物や菌類の細胞壁の構造破壊を誘導し、免疫反応成分の遊離や合成を促進させる有効な戦略である。菌類の細胞壁は主に高分子量のキチンから構成されており、ヒトの消化器官では分解が困難である。微生物発酵によって産生されるキチナーゼは、真菌の細胞壁マトリックスを破壊し、多糖類の抽出を促進するのに役立つ(Bururberg et al, 1994)。Lactiplantibacillus発酵は、胞子の細胞壁を破壊することにより、G. lucidum多糖類などの免疫活性物質の放出を効果的に促進し、これらの化合物の免疫調節能力を向上させることが可能である。より具体的には、Lp. PlantarumによるG. lucidum胞子の48時間発酵は、活性多糖構造の安定性を変えることなく、52.38%の割合で、胞子壁構造を効果的に破壊する(Nguyen and Nguyen, 2015)(Chaiyasutら, 2010)。
多糖類の免疫調節活性は、分子量、単糖組成、α/β-配置、コンフォメーション、グリコシド結合などの構造特性に大きく依存する(Chen and Huang, 2018)。多糖類は、免疫細胞の表面にあるデクチン-1、補体受容体3(CR3)、Toll様受容体(TLR)などの対応するパターン認識受容体(PRR)を認識することにより、細胞内シグナル伝達経路を活性化し、サイトカインなどの免疫分子の遺伝子発現を促進し、免疫システムを活性化することができる(Jiang et al.2010; Yin et al.2019). LABが分泌するグリコシダーゼは、多糖類の分子鎖を分解することができ、これは不溶性の高分子鎖を異なる形態を持つ小さなオリゴ糖の断片に分解することを含む。これにより、多糖類は免疫細胞の受容体タンパク質に結合し、活性化されると考えられる。LABによって発酵された多糖類は、キトサン、アルガル、マンナン、およびガラクトオリゴ糖などのいくつかの機能性オリゴ糖を生成することもあり、これらは、腸内フローラを調節し、短鎖脂肪酸の生成を促進し、腸管粘膜バリアを強化し、炎症を緩和し、腸管免疫を向上させることによって人間の腸の健康(Patel and Goyal、2011)に有益である(Cheng et al, 2019; Ma et al., 2020; Carvalho et al., 2021; Zhang N. et al., 2021)。
Streptococcus alactolyticusが介在する、ハトムギの根から得られる伝統的なハーブであるRadix astragali(RA)の発酵は、RAの総糖含量を著しく増加させる(Liang、2019年)。HPLC、HPGPC、FT-IRを用いたさらなる分析により、発酵ハトムギ多糖類(FAPS)の平均分子量は非発酵対照より低く、(1→4)-α-D-グルカンの構造を有するFAPSは強い免疫増強作用を有するようだ(Li et al.、2020)。抗酸化活性測定では、FAPSのDPPHフリーラジカル(FR)の消去率(26.40%)がAPSの消去率(20.81%)より高く、前者はシクロホスファミドによる腸管上皮細胞壁障壁機能を回復し腸管粘膜障害を解決する能力が向上することが分かった(Liang, 2019)。別の研究では、リモシラクトバチルス・ファーメンタムを介した発酵が、ロンガンパルプ多糖類(LP)の物理化学的および生物学的特性の一部を効果的に改変したことを報告した(Huang et al.、2019)。LPよりも分子量が50%低い発酵ロンガン多糖(LP-F)は、中性糖、ウロン酸、グルコースが少なく、アラビノース、ガラクトース、ラムノース、マンノースが多いことが分かった。また、LP-Fは溶解性が高く、見かけの粘度が低く、粒子径が小さく、一方で、マクロファージによる一酸化窒素(NO)とインターロイキン(IL)-6の誘導が増加し、腸内プロバイオティクスの生産が高く、強い免疫調節活性とプレバイオティクス活性を示した(Huang et al.、2019)。したがって、LABによる植物または菌類多糖の発酵修飾は、その免疫調節活性を向上させるための重要な方法である。
サポニン化合物に対する発酵の影響
ヒト腸内細菌は、糖鎖の長い分子量の大きなサポニンを、より強い薬理活性を示す分子量2以下の小さなサポニンに脱グリコシル化することが示されている。近年、サポニンの生体内変換におけるLABの役割に注目が集まっている。LABはβ-グルコシダーゼを産生することにより、前駆体サポニンを脱糖、酸化、脱水などの生化学的反応により希少な二次サポニンに変換し、生物活性を向上させることが可能であると考えられている。
Lp. plantarum M2を介した発酵の間、小腸で容易に吸収される紅参(RG)中のジンセノサイド、CK、Rh1、Rh2、Rg5、Rk1、Rg2およびRg3のレベルは67.5%増加する(Kimら、2011年)。また、発酵紅参(FRG)は、粘膜免疫系を強化することにより、in vivoでより強い抗腫瘍反応を示すことが分かっています。実際、500μg-マウス-1の用量におけるFRG阻害率は、非発酵グループよりも発酵グループで21%高かった(Kimら、2011年)。健常者の血清中の免疫グロブリンA(IgA)も、FRG投与後はベースライン時より高く(5.14 mg-mL-1)、未発酵のRGは逆の効果(-14.50 mg-mL-1)を示しました。これらの知見は、FRGが体液性免疫応答を効果的に調節できることを示しています(Kim et al.) Parkら(2014)は、Lacticaseibacillus paracasei subsp. paracasei発酵によって製造されるWoongjin発酵紅参が、非発酵RGよりも免疫関連サイトカイン産生の増加および脾細胞の増殖の刺激によって強い免疫賦活作用を有することを見出した。さらにHPLC分析により、RG抽出物は発酵後にジンセノサイド代謝物であるF2およびCKの含量を有意に増加させることが分かりました。高麗人参に天然に存在しないCKは、微生物の発酵と変換から生成される強力な新規代謝物である(Parkら、2014)。Kimら(2019)は、RG中のRd.、Rh1、F2、Rg3などのいくつかの機能性アグリコンのレベルが、Bifidobacterium animalis subsp.lactis LT 19-2で発酵することによって増加したことも報告しています。FRGはRGよりもマクロファージ活性に強い影響を与え、腫瘍壊死因子(TNF)-αおよびIL-6の高い産生によって実証された。最近の研究では、乳酸菌とビフィドバクテリウム・ブレーベによる多株発酵が、ルシデン酸A、ルシデン酸N、ガノデリック酸J、ガノデリック酸AM1などのG. lucidumトリテルペノイドをガノデリック酸C2などの有機酸に変化させることがわかった(Li et al, 2021)。この発酵生成物は、非発酵抽出物よりも粘膜バリアの免疫機能に大きな影響を与え、免疫抑制マウスの腸内細菌叢異状を逆転させた(Li et al.、2021年)。このように、LAB発酵によってサポニンの構造構成を変更することは、天然医薬品の免疫調節効果を高めるための有効な戦略である。
ポリフェノール化合物に対する発酵の効果
発酵は、植物のポリフェノール含有量を増加させる有効な手段である。発酵は、微生物酵素を用いて植物細胞壁の構造崩壊を促進することに加え、微生物の代謝経路を改変し、新規生理活性物質の遊離や合成をもたらす可能性があります(Hussain et al.、2016)。Gongら(2020a)は、Lp.plantarum SHYZ001を介した発酵がDendrobium candidumのフェノール含有量を変化させることができると報告した。HPLCベースの分析では、発酵ブロス中のフェノール酸、4-ヒドロキシ桂皮酸とシリンガットのレベルが、それぞれ42.2914 μg-g-1と12.3510 μg-1増加したことが示された。また、ノンターゲットメタボロミクス解析により、フェノール化合物の生成の主要経路であるフェニルプロパノイド代謝経路が発酵中にアップレギュレートされることがわかった(Gong et al.2020a)。さらに、Levilactobacillus brevis、Limosilactobacillus fermentum、Lp. plantarumcanなどのいくつかのLAB株は、ポリフェノールオキシダーゼ、デカルボキシラーゼ、レダクターゼなどの多様な酵素を介してフェノール酸を代謝する(Filanninoら、2015年)。Lacticaseibacillus rhamnosusとLp.plantarum株で発酵させた後、ケルセチン-3-O-ルチノシド、ヒドロキシシンナミック酸、フラボノール、アントシアニンなどのポリフェノール化合物の含有量が増え、エルダーベリージュースのジヒドロカフェ酸やカテコールなどのいくつかのフェノール代謝産物が生成します(Ricci et al.、2011)。
生薬に含まれるフラボノイドは、発酵中に構造変化を起こし、免疫調節活性を高めることができる。自然界では、バイカリン、アピゲニン、ルテオリン、イカリンのようなグリコシル化形態で頻繁に発見されます。天然のフラボノイド配糖体は、ヒトの生体に直接吸収されることはなく、まず腸内酵素または大腸の微生物叢によってアグリコンに加水分解されなければならない(Manachら、2004年)。このように、フラボノイドの生物活性は細菌の代謝に大きく依存している。発酵中、LABが産生するβ-グルコシダーゼおよび他のヒドロラーゼは、糖間結合の切断に寄与し、加水分解されてフェノールアグリコン部分を放出するグリコシドをマッチングさせると考えられる(Martinsら, 2011)。キムチから分離したLAB株であるLactiplantibacillus pentosus NGI01は、最適な生変換条件下で、フラボノイド配糖体のヘスペリジンとルチンをそれぞれ69.1%と19.4%をアグリコン形態のヘスペレチンとケルセチンに変換した(Park et al., 2021)。別の研究では、Limosilactobacillus mucosae INIA P508の形質転換株は、β-グルコシダーゼGLU913の異種生産により、リグナン、イソフラボノイド、フラボン、フラバノンの脱グリコシル化触媒を有効に行うことができた(Gaya et al.、2020年)。また、Lp.plantarum株にはエステラーゼ酵素が広く存在し、植物マトリックス中のフラボノール配糖体の代謝を促進している(Esteban-Torres et al.) Lp. plantarum株とLevilactobacillus brevis POM4を用いたカクタスペアパルプの発酵により、ケムフェロールとイソラムネチンという2種類のフラボノイド誘導体が生成され、抗酸化作用と免疫調節作用が増強された(Filannino et al.) これらの知見は、フラボノイドの構造を改変し、そのバイオアベイラビリティを向上させるのに十分なヒドロラーゼ活性を有するLABを使用することが、天然物の免疫調節活性を向上させる一つの方法であることを示すものであった。
生理活性アミノ酸に及ぼす発酵の影響
GABAを生産する仕組みとして、微生物を利用したバイオコンバージョン技術が注目されています。GABAの前駆体であるグルタミン酸は、天然植物や動物薬に広く含まれています。LAB由来のグルタミン酸脱炭酸酵素と補酵素因子である5′-ピリドキサールリン酸は、グルタミン酸をGABAに変換することができる。Kimら(2014)は,Levilactobacillus brevis GABA 100とBifidobacterium bifidum BGN4の共培養により誘導されたGastrodia elata Bl.の発酵で,大量の有機酸とGABAが生成され,グルタミン酸の75.3%がGABAに変換されたと報告している。Levilactobacillus brevis POM4を用いたPortulaca oleracea L.の発酵でも、GABAの生成量が多く(約400%)、抗栄養因子であるシュウ酸のレベルが30%低下した(Filannino et al.,2017)。また、Yooら(2022)は、Lacticaseibacillus rhamnosus LFR20-004とLatilactobacillus sakei LFR20-007による鹿角ベルベットの発酵が、シアル酸やGABAなどの生理活性アミノ酸の濃度を増大させたと報告しています。発酵鹿角は、IL-6、IL-10、IFN-γ、TNF-αの産生を促進し、強力な免疫強化活性を示したが、これはおそらくこれらの生理活性分子の増加によるものである(Yooら、2022)。
乳酸菌に対する生薬の影響
異なる天然物は、異なる方法でLABの増殖に影響を与えると考えられている。一般に、多糖類は細菌の成長、繁殖、SCFAsの生産を含む代謝過程を促進するためのエネルギー源として利用することができる。SCFAsは、様々な細菌から特定の酵素を介した解糖により生産される(Macfarlane and Macfarlane, 2003)。それらの生産は、菌株の種、増殖状態、および基質の特性などの複数の要因に関連している(Özcelikら、2016;Annunziataら、2020;Hadiniaら、2022)。試験管内での発酵は、大量のSCFAを得るための実行可能な戦略として認識されている。SCFAである酢酸、プロピオン酸、酪酸は、韓国人個体および発酵食品から分離した4つのLAB菌株によって生産され、そのうちBifidobacterium bifidum MG731は4,998.6μg-1を誘導した。 6 μg-g-1 SCFA 産生、Bifidobacterium lactis MG741 2,613.9 μg-g-1 、Ligilactobacillus salivarius MG242 1,456.1 μg-g-1 および Lp. plantarum MG989 630.2 μg-g-1 を誘導した (Kang et al., 2010), 2010). さらに、Polygonatum cyrtonema由来のフルクタンおよびガラクタンは、BifidobacteriumおよびLigilactobacillusプロバイオティクス菌株によって選択的に利用され、その成長促進特性が強調された(Zhang J. et al.、2021年)。ラムノース、アラビノース、キシロースを主成分とするPolygonatum sibiricum多糖類は、腸内微生物発酵により単糖類に分解され、酢酸、プロピオン酸、乳酸を生成することができる(Luoら、2022年)。ランの塊茎から得られる貴重な成分であるSarepは、そのグルコマンナン含有量からプレバイオティックの可能性を示し、Bifidobacterium infantisの増殖に炭素源を提供し、そのSCFA代謝物の生産を促進(1.27 g-L-1 から 1.94 g-L-1) (Usta-Gorgun and Yilmaz-Ersan, 2020)することが示された。G. lucidum多糖類は、腸管免疫細胞のGタンパク質共役型受容体43(GPR43)を活性化し、抗炎症および腫瘍免疫調節効果を引き出すことにより、大腸炎マウスの腸内のLigilactobacillus、BifidobacteriumおよびSCFAの相対存在量を有意に増加させた(Guoら、2021年)。最近の研究では、Polygonatum kingianumの多糖類がLigilactobacillus faecisのクオラムセンシングシグナルを強化し、ldh遺伝子、metE遺伝子、adh2タンパク質をアップレギュレートし、mvK遺伝子をダウンレギュレートして腸のSCFAs生成に影響するなどSCFAの合成と代謝に関する遺伝子やタンパク質の発現を調節することがわかった(Yang et al.) ハーブ成分がLABの代謝活性に与える影響に関する研究の多くは、SCFAに着目しています。一方、EPSやバクテリオシンなど、漢方薬に含まれる他の免疫活性成分の効果については、まだ解明されていない。
乳酸菌発酵生薬の免疫調節機構
乳酸菌発酵生薬は、主に自然免疫反応の活性化、適応免疫反応の制御、腸管免疫恒常性の維持により、ヒトの免疫機能を変化させる。表1に、様々な動物あるいは細胞モデルにおける免疫調節作用とその分子機構の概要を示す(表1)。
自然免疫系の活性化
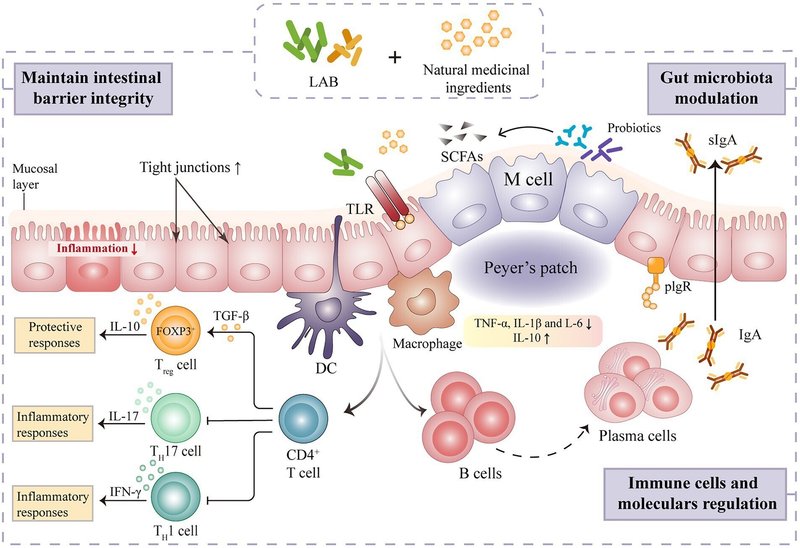
自然免疫系の構成要素(その組織バリアー、自然免疫細胞およびタンパク質など)は、病原体の検出とクリアランスの第一線として機能する。天然薬物のLAB発酵は、自然免疫細胞の活性化、免疫関連サイトカインの調節、補体系の活性化により、免疫力を向上させることができる(図2)。
図2
www.frontiersin.org
図2.自然免疫系と適応免疫系の仕組み 乳酸菌で発酵させた天然医薬品の自然免疫と適応免疫の調節機構。
マクロファージ
マクロファージは自然免疫細胞の一種で、恒常性の維持や外来病原体への抵抗に重要な役割を担っています(Lee et al.、2020)。これらの細胞は、外来病原体や腫瘍細胞を直接排除する免疫エフェクターとして働くとともに、不可欠な抗原提示細胞(APC)であり、炎症促進および抗炎症サイトカインやケモカインを分泌して免疫応答を活性化します(Aminin and Wang, 2021)。マクロファージのFcγ受容体(FcγR)は、これらの細胞が外来抗原と接触する場所で凝集することにより、貪食を促進する。LAB発酵は、FcγRおよびTLR2シグナルを活性化し(Kwonら、2018)、マクロファージに自然免疫制御に関与するサイトカインであるTNF-αおよびIL-6の産生を誘導することによって貪食を促進できる(Wang and He, 2020)。Kimら(2019)は、Bifidobacterium animalis subsp. lactisによる紅参発酵後、マイトジェン活性化プロテインキナーゼ(MAPK)およびNF-κBシグナルによりマクロファージ活性が著しく上昇したと報告しています。紅参発酵は、TNF-αやIL-6の産生を促進することで、脾臓細胞や骨髄由来マクロファージ(BMDM)などの中枢性免疫細胞も活性化させた。マクロファージから分泌されるNOも免疫細胞の活性化を誘導し、病原体に対する抵抗力を促進します(Huら、2021年)。Moら(2021)は、3%RGE添加培地で培養したLacticaseibacillus paracasei HY7017がNO産生を劇的に高めることを観察した。また、HY7017-RGEsで処理すると、NO合成の重要な酵素であるiNOSとCOX-2のmRNAレベルが高くなった。この過程では、抗炎症サイトカインであるIL-10の分泌には影響がなかったことから、発酵紅参は過剰な炎症を起こすことなく免疫反応を高めることが示された(Moら、2021年)。
樹状細胞
樹状細胞(DC)は、免疫センチネルとして機能し、免疫反応の開始を助けるプロフェッショナルAPCの一種です(Zannaら、2021)。未熟なDCは強い移動能力を持ち、成熟したDCはナイーブT細胞を効果的に活性化し、免疫応答の開始、調節、維持に関与する(Zhou and Ye, 2021)。成熟DCは、高レベルのNO、IL12、およびケモカイン、RANTESを合成し、CD86および主要組織適合性複合体(MHC)などの共刺激分子を発現し、完全なT細胞活性化を誘導する(Zhuら、2019年)。いくつかの研究は、LAB発酵生薬が、おそらくMAPK-JNK、p38 MAPK、およびNF-κBシグナル伝達経路の活性化を通じて、DC成熟を促進することによって免疫を調節することを示している(Zhangら、2009年)。マウスモデルにおいて、FAPSはDC成熟関連シグナル伝達に影響を与え、それによってその成熟を促進することが示された(Bian, 2017)。その結果、異なる用量のFAPS(50、100、200μg-mL-1)は、骨髄由来のDCを誘導し、未暴露のDCよりも高いレベルのIL-12、TNF-α、IL-1β、IL-6の生成および関連遺伝子発現を引き起こすことが明らかにされた。DCを活性化することで、FAPSは6.8~19%高いT細胞増殖を促進することも示されました(Bian, 2017)。別の研究では、発酵生薬で処理したDCは、B細胞の増殖と分化を直接刺激できることがわかりました。Zhangら(2009)は、発酵ノニ滲出液(FNE)が成熟DCおよび混合脾臓細胞培養Bリンパ球の増殖を促進したことを報告し、FNEが骨髄由来のDC成熟を誘導し、B細胞分化および免疫グロブリンクラス転換を促進するために必要なDC誘導免疫応答を刺激できることを示している。
ナチュラルキラー細胞
自然免疫系のエフェクターリンパ球であるナチュラルキラー(NK)細胞は、MHCクラスIを欠く、あるいはNK細胞受容体特異的リガンドを過剰発現する異常細胞をパトロールすることにより、腫瘍および微生物感染を抑制する(Lanier, 2005; Vivier et al.) これらの細胞を認識した後、活性化されたNK細胞は、標的細胞を殺すためにグランザイムとパーフォリンを放出し、免疫調節を助けるIFN-γ、TNF-α、IL-10などのいくつかのサイトカインを分泌する(Kim et al.、2007)。自己免疫、移植およびウイルス感染に関する最近の研究では、NK細胞をT細胞媒介応答の制御と関連付けている(Zwirnerら、2021年)。LABとその発酵製品は、NK細胞の活性と細胞傷害性を高めることが示されている。ある研究では、加熱殺菌したラクトバチルス・アシドフィルスLa205が、顆粒エキソサイトーシスを増加させることにより、NK細胞の細胞毒性を直接刺激できることが分かった(Cheonら、2011年)。Kang ら(2010)は、マウスリンパ腫細胞(Yac-1)に対する細胞毒性影響を測定することにより、発酵した高麗人参エキスの NK 細胞活性への影響を評価しました。この抽出物は、用量依存的にNK細胞の細胞傷害性を高め、IFN-γの分泌を促進することが示されました(Kang et al.、2010)。
補体システム
補体系には、約40種類の血清タンパク質からなる酵素反応のカスケードが存在する。補体活性化経路には、古典的経路、代替経路、レクチン経路の3つがある(Garredら、2021年)。代替経路はバイパス経路とも呼ばれ、微生物や異物によって直接C3が活性化され、抗体に依存しない。補体成分であるB、D、P因子は、C3インベルターゼとC5インベルターゼの産生、膜攻撃複合体(MAC)の形成、標的細胞の破壊につながる酵素触媒による補体反応経路のカスケードの誘導に役立つ(Gupta and Tripathy, 2020)。LAB-発酵によって生成された断片化された多糖類分子は、代替経路を通じて補体を活性化し、免疫応答を制御することが示されている(Watanabe et al., 2015; Mastuba et al., 2017; Chen et al., 2022)。
適応免疫系の制御
自然免疫系は、生殖細胞にコードされた限られた数のPRRを用いて、一定の病原体関連分子パターン(PAMPs)を認識する。対照的に、適応免疫系は、TおよびBリンパ球上の複数の抗原受容体の産生、およびその後の適切な抗原特異的受容体を有するそれらの細胞の活性化とクローニングに依存している(Schenten and Medzhitov, 2011)。LAB発酵天然薬物中の活性分子は、体液性および細胞性免疫機能を変化させることにより、免疫反応を調節することができます(図2)。
体液性免疫
体液性免疫とは、Bリンパ球が産生する抗体を介した適応的な免疫防御機構を指す。抗原刺激により、B細胞は抗体産生B細胞株や形質細胞に変化し、抗原に特異的に結合する抗体、すなわち免疫グロブリン(Ig)を産生するようになる。IgGは体液性免疫反応の必須成分であり、ヒト血清中に存在する主要なIgクラスである。IgGがマクロファージ、好中球、NK細胞の表面にあるFc受容体に結合すると、抗体依存性細胞傷害(ADCC)が誘導される(Dixonら、2021年)。IgAは、ヒトに存在する最も豊富な免疫グロブリンであり、血清型と分泌型(sIgA)の2つの形態で存在する。IgAは、主にIgA Fc受容体I(FcRI)との相互作用により、粘膜の恒常性維持に関与しています(Mkaddemら、2014年)。IgMは、結合鎖ポリペプチドを含む5量体で、感染後に最初に産生される抗体であり、しばしば免疫防御の第一線とみなされる(Hiramoto et al.) 紅参発酵は、多糖類やサポニンなどの免疫反応性成分の存在に潜在的に起因して、健康なヒト血清中のIgGおよびIgAレベルを増加させることが示されている(Kimら、2011年)。ある研究では、Lp. PlantarumによるGynostemma pentaphyllum経口液の発酵が、免疫抑制マウスの血清中へのIgA、IgG、IgMおよび関連サイトカインの分泌を促進し、その免疫力を高めることを発見した(Du、2021年)。これらの知見は、LAB発酵生薬が抗原特異的な免疫反応を高めることができることを示している。
細胞媒介性免疫(Cell-mediated immunity
細胞媒介性免疫とは、抗体とは無関係に、抗原特異的なT細胞が細胞内病原体を認識して駆除する能力に依存する免疫反応です。未熟なT細胞は、同種の抗原で刺激されると増殖し、エフェクター細胞に分化した後、アポトーシスを起こすか、長寿命の記憶細胞の前駆体として生き残る。IL-2は、T細胞の成長、増殖、およびその後の分化を刺激し、その細胞傷害性効果を高める強力なT細胞成長因子である(Malek、2003;Rochmanら、2009)。活性化されたT細胞は、TNFスーパーファミリーのメンバーであり、T細胞の応答性を制御するコミュニケーションシステムにおいて重要な役割を果たすTNFSF14を発現しています(Wang and Fu, 2004)。Kimら(2015)は、高麗人参の芽をLp.plantarumとLacticaseibacillus caseiで発酵させ、T細胞活性化に対する混合発酵ブロスの効果を研究しました。このブロスは、T細胞においてIL-1βの発現を防ぎ、IL-2およびTNFSF14の発現を高めることによって炎症を抑制することが示されました(Kim et al.) CD4+T細胞には、Tヘルパータイプ1(Th1)と制御性T細胞(Treg)の2種類があります。Th1細胞はIFN-γとIL-2を産生し、細胞媒介性免疫を誘導するが(Murakami et al., 2004)、Treg細胞は自己寛容と免疫細胞の恒常性の維持に不可欠である(Göschl et al., 2019)。RG発酵産物は、脾臓リンパ系T細胞のTh1細胞およびTreg細胞への分化を促進し、免疫抑制を軽減することができる(Kimら、2018年)。CD4+/CD8+比は、免疫機能の重要な指標である。Lp. plantarum NCU137-発酵のCoix seedは、脾臓指数を有意に増加させ、脾臓Tリンパ球サブセットのCD4+/CD8+比を回復し、TNF- αおよびIL-6の産生を低下させ、炎症を抑えることがわかり、その効果は未発酵群よりも良好でした(Wang et al. 2022)。G. lucidum子実体の混合プロバイオティクス(Lactobacillus acidophilusとBifidobacterium breve)仲介発酵は、マウス脾臓のCD4+ TおよびCD8+ Tリンパ球の割合を増加させ、T細胞の形質転換を改善した(Li et al., 2021)。
腸管免疫ホメオスタシスの維持
腸管は主に上皮細胞と免疫細胞から構成されており、人体で最大の免疫器官となっています(Mahapatro et al.、2021)。腸管免疫とホメオスタシスは、バランスのとれた腸内細菌叢に依存しています。異なる腸管免疫細胞のタイプが微生物のコロニー形成の影響を受け、腸管免疫細胞は直接的および間接的な方法で微生物集団に影響を与えることができます。LABは、ムチンMUC2、トレフォイル因子3、レジスチン様分子β3などの杯細胞関連遺伝子を調節することにより、腸管バリアに影響を与えると考えられています(Renら、2018)。また、乳酸発酵は、腸内微生物の領域構造を調節し、腸管免疫細胞や因子を活性化し、炎症を緩和するという顕著な役割を果たすことが示されている(図3)。FAPSは、免疫抑制マウスの免疫指標と腸の形態を回復させることができました(Liang, 2019)。実際、sIgA、α鎖および高分子免疫グロブリン受容体(pIgR)のmRNA発現は、食餌FAPS補給後にすべて有意にアップレギュレートされた。FAPSはまた、小腸のタイトジャンクション(TJ)および接着接合部(AJ)関連タンパク質の発現を促進し、腸管上皮バリアの完全性を維持することができます(Liang、2019)。最近の研究では、発酵人参(390 mg-kg-1日-1)の補給が、TLR4/MAPKシグナルを活性化し、腸のバリアを回復させることによって、マウスのリポポリサッカライド誘発性炎症を有意に改善することが示されました(Fan et al.、2021年)。さらに、天然薬のLAB媒介発酵の免疫調節活性は、腸のパイエル板におけるCD4+ T細胞増殖の直接調節と正の相関がある(Liら、2021)。Gongら、(2020a)は、Lp.plantarum SHY001発酵Dendrobium candidum(FDC)の、ゼブラフィッシュのオキサゾロン誘発腸炎症モデルにおける腸内細菌多様性と腸管免疫調節への影響を評価した。FDC(0.15-g L-1)処理後、Lactobacillus、Faecalibacterium、Rummeliibacillusなどの有益な細菌の存在量が増加し、ShewanellaやMycobacteriumなどの腸の日和見病原体のレベルが低下しました(Gong et al.) また、FDCは腸管SCFAや粘膜バリア関連タンパク質であるMUC2、オクルディン、ZO-1の産生を有意に促進し、炎症性サイトカインであるIL-4、IL-10、TNF-αの分泌を減少させた(Gong et al.、2020a)。これらの知見は、FDCが腸内細菌叢とその代謝物を調節することによって腸の健康を増進し、腸の炎症を緩和し、宿主免疫系を調節することができることを示すものである。
図3
www.frontiersin.org
図3. 乳酸菌で発酵させた天然医薬品の腸管免疫調節のメカニズム。
まとめ
漢方薬には、多糖類、サポニン、フラボノイド、フェノール酸などの免疫調節活性成分があり、免疫機能の調節に重要な役割を担っている。したがって、これらの成分のバイオアベイラビリティは、ハーブの免疫調節効果にとって不可欠である。最近の研究では、腸内細菌が薬用活性物質の代謝・吸収に大きな影響を与えることが分かっています(Wang S. et al., 2019; Gong et al., 2020b; Zhao et al., 2022)。しかし、免疫機能の乱れは、薬物吸収に悪影響を及ぼす腸内細菌叢のアンバランスを伴うことが多く、このような腸内細菌叢のアンバランスは、薬物吸収に悪影響を及ぼす。試験管内での生薬のLAB発酵は、薬物の吸収と代謝を補助する効果的な戦略として利用されている。LABは、3つの方法で発酵生薬の免疫調節活性を高める上で重要な役割を果たした。第一に、LABは発酵の過程でSCFAsなどの免疫調節活性物質を産生することができる。第二に、LABによる発酵は、生薬から本来の免疫活性物質の放出を促進する。第三に、LABは、これらの薬物の特定の成分の構造改変を通じて、新規の免疫調節化合物の産生を誘導する可能性がある。
乳酸菌は発酵食品、植物、動物、ヒトに広く分布しているが、すべての菌株が生薬の発酵に使用できるわけではない。一般に、植物由来の菌株は、ポリフェノールなどの植物成分に対する耐性が高いため、スターターカルチャーとしての使用に最も適している(Lee and Paik, 2017)。食品発酵と比較して、生薬発酵に適した理想的な工業用スターターはまだ研究段階である。現在、生薬発酵研究に用いられている菌株の多くは、発酵食品や腸内フローラ由来のものであり、生薬発酵により適した薬用植物由来の内生菌や外生菌は十分に研究・開発されていないものもあります。実際、内生微生物が宿主の薬用植物の二次代謝を誘導することはよくあることである。薬用植物微生物が宿主植物のアルカロイド、ステロイド、テルペノイド、その他の重要な薬効成分の収量に影響を与えることが研究で示されている(Pangら、2021年)。これらの微生物は、重要な機能性二次代謝産物を合成する重要な遺伝子や酵素を含んでいる可能性がある。したがって、薬用植物から内生あるいは外生LABを分離・スクリーニングすることは、薬草の効率的な発酵に不可欠であろう。LABの活性は、種と株の両方に特異的であることは注目に値する。また、LABの代謝活性は、発酵基質(ハーブの種類)や培養条件(時間、温度、pH、栄養添加など)といった外的要因にも影響される。漢方薬発酵に使用する培養スターターをスクリーニングする際には、LABのこれらの特性を考慮する必要がある。
LAB発酵生薬の免疫調節活性に関する現在の研究は、生薬成分の役割にのみ焦点が当てられており、LAB、特に生菌の免疫調節特性は考慮されていない。LAB発酵ハーブの経口投与後、LAB微生物は腸内に定着し、EPS、SCFA、バクテリオシンなどの免疫調節活性代謝産物を分泌し続け、免疫系に強く持続的な影響を与えることが可能である。
LABがハーブ成分を変化させる、あるいは構造的に変化させるメカニズムについては、包括的かつ体系的に研究されていない。加水分解が主な研究対象であり、脱メチル化反応や酸化還元反応はほとんど報告されていない。メタボロミクス、トランスクリプトミクス、その他のマルチオミクス技術は、微生物発酵生薬の生体内変換機構を探るために徐々に適用されつつあります。しかし、高い活性を持つ新規代謝物の同定や、その後の免疫調節における役割の証明はまだ十分ではありません。つまり、微生物発酵生薬と生薬の複雑な相互作用は、免疫調節成分の発見とそれらが免疫反応を調節するメカニズムの解明という、新たな機会と課題を提供するものである。
著者協力
HZ、LG、DY、XD が文献を検討し、原稿を執筆した。HZは要約と図表の作成を行った。提出された原稿は全著者によって承認された。
利益相反
著者らは、本研究が利益相反の可能性があると解釈される商業的または金銭的関係のない状態で実施されたことを宣言する。
出版社からのコメント
本論文で述べられたすべての主張は、著者個人のものであり、必ずしも所属団体、出版社、編集者、査読者のものを代表するものではありません。この記事で評価される可能性のある製品,あるいはそのメーカーによる主張は,出版社によって保証または承認されたものではありません.
参考文献
Abbas, M., Saeed, F., Anjum, F. M., Afzaal, M., Tufail, T., Bashir, M. S., et al. (2017). 天然ポリフェノール:概説。Int. J. Food Prop. 20, 1689-1699. doi: 10.1080/10942912.2016.1220393.
クロスレフフルテキスト|Google Scholar
Abdou, A. M., Higashiguchi, S., Horie, K., Kim, M., Hatta, H., and Yokogoshi, H. (2006). ヒトにおけるγ-アミノ酪酸(GABA)投与によるリラックス効果および免疫力向上効果. バイオファクターズ 26, 201-208.
PubMed Abstract|CrossRef Full Text|Google Scholar
Alesci, A., Nicosia, N., Fumia, A., Giorgianni, F., Santini, A., and Cicero, N. (2022). レスベラトロールと免疫細胞:ヒトの健康増進へのつながり。モレキュールズ 27:424.
PubMed Abstract|CrossRef Full Text|Google Scholar
Aminin, D., and Wang, Y. M. (2021). 抗がん剤細胞免疫療法における「武器」としてのマクロファージ。KJMS 37, 749-758. doi: 10.1002/kjm2.12405.
PubMed Abstract|CrossRef Full Text|Google Scholar
Annunziata, G., Arnone, A., Ciampaglia, R., Tenore, G. C., and Novellino, E. (2020).食品および飲料の発酵。短鎖脂肪酸に焦点を当てた生物活性化合物の利用率を高めるためのツールとしての食品および飲料の発酵。Foods. 9:999. doi: 10.3390/foods9080999
PubMed Abstract|CrossRef Full Text|Google Scholar
ビアン、Y. B. (2017). 発酵ハトムギ多糖類の調製とin vitroにおけるマウス骨髄由来樹状細胞成熟のシグナル伝達経路に及ぼす影響。[学位論文/修士論文]. [北京(中国)]。中国農業科学院
Google Scholar
Bjurstöm, H., Wang, J., Ericsson, I., Bengtsson, M., Liu, Y., Kumar-Mendu, S., et al.(2008)。GABA, a natural immunomodulator of T lymphocytes. J. Neuroimmunol. 205, 44-50. doi: 10.1016/j.jneuroim.2008.08.017.
PubMed Abstract | CrossRef Full Text | Google Scholar
Brurberg, M. B., Haandrikman, A. J., Leenhouts, K. J., Venema, G., and Nes, I. F. (1994).キチン分解酵素の発現。セラティア・マルセセンス由来キチナーゼ遺伝子のラクトコッカス・ラクティスおよびラクトバチルス・プランタラムにおける発現。Appl. Microbiol. Biotechnol. 42, 108-115.
PubMed Abstract|CrossRef Full Text|Google Scholar
Carvalho, P., Andrade, M., Trindade, L. M., Leocádio, P., Alvarez-Leite, J. I., Dos Reis, D. C., et al.(2021)。フラクトオリゴ糖を用いた予防的・治療的サプリメントは5-フルオロウラシルによる粘膜炎後の腸のホメオスタシスを改善する。Biomed. Pharmacother. 133:111012.doi: 10.1016/j.biopha.2020.111012
PubMed Abstract|CrossRef Full Text|Google Scholar
チャイヤスット、C.、クルアタマ、C.、シリルン、S. (2010). を用いた発酵によるGanoderma lucidumの胞子の破砕。Afr. J. Biotechnol. 9, 7379-7382. doi: 10.5897/AJB09.1085.
クロスレフフルテキスト|Google Scholar
チャップリン、D.免疫反応の概要 J. Allergy Clin. Immunol. 125, S3-S23. doi: 10.1016/j.jaci.2009.12.980.
PubMed Abstract|CrossRef Full Text|Google Scholar
Chen, F., and Huang, G. (2018). 多糖類およびその誘導体の調製と免疫学的活性。Int. J. Biol. Macromol. 112, 211-216. doi: 10.1016/j.ijbiomac.2018.01.169.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Chen, Q., Sun, S., Mei, C., Zhao, J., Zhang, H., Wang, G., et al.(2022年)。生物結合能力、抗酸化能力、腸内環境修復能力が乳酸菌のペルフルオロオクタンスルホン酸毒性緩和能力を共同で決定している。Environ. 166:107388.論文番号: 10.1016/j.envint.2022.107388
PubMed Abstract|CrossRef Full Text|Google Scholar
Cheng, Y. F., Chen, Y. P., Chen, R., Su, Y., Zhang, R. Q., He, Q. F., et al. (2019). Dietary mannan oligosaccharide ameliorates cyclic heat stress-induced damage on intestinal oxidative status and barrier integrity of broilers(食餌性マンナンオリゴ糖はブロイラーにおける周期的熱ストレス誘発性ダメージを改善する。Poult. Sci. 98, 4767-4776. doi: 10.3382/ps/pez192
PubMed Abstract | CrossRef Full Text | Google Scholar
Cheon, S., Lee, K. W., Kim, K. E., Park, J. K., Park, S., Kim, C. H.、他 (2011). 加熱殺菌した乳酸菌 La205 は、顆粒エキソサイトーシスの増加を通じて NK 細胞の細胞傷害性を高める。Immunol. Lett. 136, 171-176.
PubMed Abstract|CrossRef Full Text|Google Scholar
Ciszek-Lenda, M., Nowak, B., Sróttek, M., Gamian, A., and Marcinkiewicz, J. (2011). Lactobacillus rhamnosus KL37由来エキソポリサッカライドの免疫調節能:マウスマクロファージによる炎症性メディエーターの産生に対する影響。Int. J. Exp. Pathol. 92, 382-391. doi: 10.1111/j.1365-2613.2011.00788.x
PubMed Abstract | CrossRef Full Text | Google Scholar
D'Arcy, M. S. (2022). 新生物細胞におけるオートファジーおよびアポトーシスの誘導物質として、また非新生物細胞における細胞保護物質としての生物学的活性フラボノイドの総説. セルバイオ.Int. 46, 1179-1195.
PubMed Abstract|CrossRef Full Text|Google Scholar
Dixon, K. J., Wu, J., and Walcheck, B. (2021). ヒトNK細胞によるADCCを強化するための抗腫瘍モノクローナル抗体およびfcレセプターのエンジニアリング。キャンサーズ. 13:312. doi: 10.3390/cancers13020312
PubMed Abstract|CrossRef Full Text|Google Scholar
Du, X. H. (2021). Gynostenmma Pentaphyllum発酵口腔液の調製とマウスの免疫機能への影響。[学位論文/修士論文]。[重慶(中国)].西南大学
Google Scholar
Esteban-Torres, M., Reverón, I., Mancheño, J. M., de Las Rivas, B., and Muñoz, R. (2013). ラクトバチルス・プランタラム由来のフェルロイル・エステラーゼの特性評価。Appl. Environ. Microbiol. 79, 5130-5136. doi: 10.1128/AEM.01523-13
PubMed Abstract|CrossRef Full Text|Google Scholar
範, J. J., 劉, S. T., 愛, Z. Y., 陳, Y. Y., 王, Y. H., 李, Y. B., 他 (2021). 発酵人参は、TLR4/MAPKシグナル経路の活性化と腸管バリアの改善により、リポポリサッカライド誘発の炎症反応を減衰させる。Food Funct. 12, 852-861.
PubMed Abstract|CrossRef Full Text|Google Scholar
Filannino, P., Bai, Y., Di Cagno, R., Gobbetti, M., and Gänzle, M. G. (2015). チェリージュースとブロッコリーピューレの発酵中の乳酸菌によるフェノール化合物の代謝。Food Microbiol. 46, 272-279. doi: 10.1016/j.fm.2014.08.018.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Filannino, P., Cavoski, I., Thlien, N., Vincentini, O., De Angelis, M., Silano, M.、他 (2016). カクタスクラドード(Opuntia ficus-indica L.)の乳酸発酵により、抗酸化および抗炎症作用を有するフラボノイド誘導体が生成される。PLoS One 11:e0152575. doi: 10.1371/journal.pone.0152575.
PubMed Abstract|CrossRef Full Text|Google Scholar
Filannino, P., Di Cagno, R., Trani, A., Cantatore, V., Gambacorta, G., and Gobbetti, M. (2017). 乳酸発酵は、生物起源化合物のプロファイルを豊かにし、コモンパースレイン(Portulaca oleracea L.)の機能的特徴を強化する。J. Funct. Foods 39, 175-185. doi: 10.1016/j.jff.2017.10.022.
クロスレフフルテキスト|Google Scholar
Forouzani-Haghighi、B.、Rezvani、A.、およびVazin、A.(2022)。COVID-19感染症に対する免疫標的治療:ナラティブレビュー。Iran. J. Med. Sci. 47, 291-299. doi: 10.30476/IJMS.2021.91614.2277.
PubMed Abstract|CrossRef Full Text|Google Scholar
Garred, P., Tenner, A. J., and Mollnes, T. E. (2021). 補体系の治療標的化:希少疾患からパンデミックまで。Pharmacol. 73, 792-827. doi: 10.1124/pharmrev.120.000072
PubMed Abstract|CrossRef Full Text|Google Scholar
Gaya, P., Peirotén, Á., and Landete, J. M. (2020).(ガヤ、P.、ペイロテン、およびランデテ、J.M.)。バイオテクノロジーに関心のある細菌におけるβ-グルコシダーゼの発現は、植物性食品中のリグナンおよびフラボノイドを脱グリコシル化する能力を彼らに付与する。Appl. Microbiol. Biotechnol. 104, 4903-4913.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Geraldi, A., Cui, C. H., Nguyen, T. T., and Kim, S. C. (2020). インドネシアからの細菌分離の組換えβ-グルコシダーゼによるジンセノサイドRb1の酵素的生体内変換(Enzymatic biotransformation of ginsenoside Rb1 by recombinant β-glucosidase of bacterial isolates from Indonesia. Biocatal. Agric. Biotechnol. 23:101449. doi: 10.1016/j.bcab.2019.101449.
クロスレフフルテキスト|Google Scholar
Ginwala, R., Bhavsar, R., Chigbu, D. I., Jain, P., and Khan, Z. K. (2019). アピゲニンの抗炎症活性を中心とした慢性炎症性疾患の治療におけるフラボノイドの潜在的な役割. アンチオキシダント. 8:35. doi: 10.3390/antiox8020035
PubMed Abstract|CrossRef Full Text|Google Scholar
Gong, X., Jiang, S., Tian, H., Xiang, D., and Zhang, J. (2020a). Dendrobium candidumの発酵液に含まれるポリフェノールは、腸内細菌を介した免疫反応により、ゼブラフィッシュの腸の炎症を緩和させる。Front. Immunol. 11:1542. doi: 10.3389/fimmu.2020.01542
PubMed Abstract|CrossRef Full Text|Google Scholar
Gong, X., Li, X., Bo, A., Shi, R. Y., Li, Q. Y., Lei, L. J., et al (2020b). 腸内細菌叢と漢方薬の生理活性成分との相互作用:レビュー。Pharmacol. Res. 157:104824. doi: 10.1016/j.phrs.2020.104824.
PubMed Abstract|CrossRef Full Text|Google Scholar
Göschl, L., Scheinecker, C., and Bonelli, M. (2019). 自己免疫におけるTreg細胞:同定からTregベースの治療法まで。Semin. Immunopathol. 41, 301-314. doi: 10.1007/s00281-019-00741-8
PubMed Abstract|CrossRef Full Text|Google Scholar
Guo, C., Guo, D., Fang, L., Sang, T., Wu, J., Guo, C., et al.(2021)。霊芝多糖は腸内細菌叢と免疫細胞機能を調節し、大腸の炎症と腫瘍形成を抑制する。Carbohydr. Polym. 267:118231。doi: 10.1016/j.carbpol.2021.118231
PubMed Abstract|CrossRef Full Text|Google Scholar
Guo, R., Guo, S., Gao, X., Wang, H., Hu, W., Duan, R.、他 (2020). 古代中国の漢方混合物である唐桂布袋湯を乳酸菌とともに発酵させると、漢方製剤の抗糖尿病機能が増強される。Chin. Med. 15:98. doi: 10.1186/s13020-020-00379-x
PubMed Abstract|CrossRef Full Text|Google Scholar
グプタ、P.およびトリパシー、A.S. (2020). 補体活性化の代替経路は、チャンディプラウイルス感染に対して有益な役割を持つ。Med. Microbiol. Immunol. 209, 109-124. doi: 10.1007/s00430-019-00648-z
PubMed Abstract|CrossRef Full Text|Google Scholar
Hadinia, N., Dovom, M. R. E., and Yavarmanesh, M. (2022). 短鎖脂肪酸を生産するための乳酸菌株を用いた発酵条件(温度、塩濃度、pH)の影響。LWT 165:113709. doi: 10.1016/j.lwt.2022.113709.
クロスレフフルテキスト|Google Scholar
平本 英夫・堤 敦・鈴木 亮・松岡 聡・荒井 聡・吉川 正也・他(2018).IgM五量体は、AIMタンパク質と結合する開溝を持つ非対称五角形である。サイエンス・アドバンス4:eaau1199. doi: 10.1126/sciadv.aau1199
PubMed Abstract|CrossRef Full Text|Google Scholar
Hu, J., Ma, L., Nie, Y., Chen, J., Zheng, W., Wang, X., et al. (2018). A microbiota-derived Bacteriocin targets the host to confer diarrhea resistance in early-weeding piglets. セルホストマイクロベ 24, 817-832.e8. doi: 10.1016/j.chom.2018.11.006
PubMed Abstract|CrossRef Full Text|Google Scholar
Hu, Q., Shi, J., Zhang, J., Wang, Y., Guo, Y., and Zhang, Z. (2021). 一酸化窒素が介在する免疫・免疫療法における制御機能の進展と展望. Adv. Ther. 4:2100032. doi: 10.1002/adtp.202100032.
クロスレフフルテキスト|Google Scholar
Huang, F., Hong, R., Zhang, R., Yi, Y., Dong, L., Liu, L., et al.(2019)。乳酸菌発酵によって改質された竜眼パルプ多糖の物理化学的および生物学的特性。Int. J. Biol. Macromol. 125, 232-237. doi: 10.1016/j.ijbiomac.2018.12.061
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Huang, F., Teng, K., Liu, Y., Cao, Y., Wang, T., Ma, C., et al. (2021). バクテリオシン:ヒトの健康への可能性。Oxid. Med. セル. Longev. 2021, 5518825-5518817.
PubMed Abstract|CrossRef Full Text|Google Scholar
Huang, N., Yu, D., Huo, J., Wu, J., Chen, Y., Du, X., et al.(2022)。Dioscorea nipponicaの内生菌による生変換後のサポニン成分の研究 C39. J. Anal. Methods Chem. 2022, 2943177-2943115. doi: 10.1155/2022/2943177
PubMed Abstract|CrossRef Full Text|Google Scholar
Hussain, A., Bose, S., Wang, J. H., Yadav, M. K., Mahajan, G. B., and Kim, H. (2016). 発酵、生薬の生物活性を高めるための実現可能な戦略。Food Res. Int. 81, 1-16. doi: 10.1016/j.foodres.2015.12.026.
クロスレフフルテキスト|Google Scholar
Iraporda, C., Errea, A., Romanin, D. E., Cayet, D., Pereyra, E., Pignataro, O., et al.(2015年)。微生物発酵によって生成された乳酸と短鎖脂肪酸は、腸管上皮細胞および骨髄系細胞における炎症性反応をダウンレギュレートする。イムノバイオロジー 220, 1161-1169. doi: 10.1016/j.imbio.2015.06.004
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Jiang, M. H., Zhu, L., and Jiang, J. G. (2010). 漢方薬由来の多糖類の免疫調節作用。Expert Opin. Ther. Targets 14, 1367-1402. doi: 10.1517/14728222.2010.531010
PubMed Abstract|CrossRef Full Text|Google Scholar
Kang, D. Z., Liu, D., Jo, S. Y., Kang, T. B., Lee, J. M., Suh, H. J., 他 (2010). 発酵した高麗人参抽出物のマウスにおける腫瘍転移に対する効果。Food Sci. Biotechnol. 19, 1559-1565. doi: 10.1007/s10068-010-0221-9
クロスレフフルテキスト|Google Scholar
Keyt, B. A., Baliga, R., Sinclair, A. M., Carroll, S. F., and Peterson, M. S. (2020). IgM抗体の構造、機能、および治療上の使用。アンティボディーズ 9:53.
PubMed Abstract|CrossRef Full Text|Google Scholar
Khan, S. A., Liu, L., Lai, T., Zhang, R., Wei, Z., Xiao, J., et al. (2018). 乳酸菌で発酵させた乾燥リュウガンのフェノールプロファイル、遊離アミノ酸組成および抗酸化能。J. Food Sci. Technol. 55, 4782-4791. doi: 10.1007/s13197-018-3411-8
PubMed Abstract|CrossRef Full Text|Google Scholar
Kim, J. H., Doo, E. H., Jeong, M., Kim, S., Lee, Y. Y., Yang, J., et al. (2019). Bifidobacterium animalis Subsp. lactis LT 19-2を用いた発酵による紅参の免疫調節機能の向上。ニュートリエンツ11:1481. doi: 10.3390/nu11071481
PubMed Abstract|CrossRef Full Text|Google Scholar
Kim, S. H., Hong, J. H., Lee, J. E., and Lee, Y. C. (2017). カンゾウ根の主要生理活性成分である18β-グリチルレチン酸は、喘息マウスモデルにおいてTh2サイトカイン、GATA-3、STAT6、Foxp3転写因子の調節により気道炎症を減弱させる。Environ. Toxicol. Pharmacol. 52, 99-113. doi: 10.1016/j.etap.2017.03.011
PubMed Abstract|CrossRef Full Text|Google Scholar
Kim, J. K., Kim, J. Y., Jang, S. E., Choi, M. S., Jang, H. M., Yoo, H. H., et al. (2018). 発酵紅参はマクロファージ活性化およびT細胞分化を制御することにより、シクロホスファミド誘発免疫抑制および2,4,6-トリニトロベンゼンスルホン酸誘発大腸炎をマウスで緩和する。Am. J. Chin. Med. 46, 1879-1897. doi: 10.1142/S0192415X18500945
クロスレフフルテキスト|Google Scholar
Kim, K. H., Kim, S. H., and Yook, H. S. (2015). 様々な微生物によって発酵させたPanax ginsengの花蕾がT細胞免疫応答の活性化に及ぼす影響。Food Sci. Biotechnol. 24, 1061-1067. doi: 10.1007/s10068-015-0136-6
クロスレフフルテキスト|Google Scholar
キム、T. W. 、イ、T. Y. 、ベ、H. C. 、ハーム、J. H. 、キム、Y. H. 、パク、C. 、その他(2007)。高分子量ポリガンマグルタミン酸の経口投与は、NK細胞を介した抗腫瘍免疫を誘導する。J. Immunol. 179, 775-780.
PubMed Abstract|CrossRef Full Text|Google Scholar
Kim, J. A., Park, M. S., Kang, S. A., and Ji, G. E. (2014). Gastrodia elata Bl.の発酵時におけるγ-アミノ酪酸の生産(Bifidobacterium bifidum BGN4とLactobacillus brevis GABA 100の共培養により)。食品科学, バイオテクノロジー. 23, 459-466. doi: 10.1007/s10068-014-0063-y
クロスレフフルテキスト|Google Scholar
キム・ビー・ジー、シン・ケイ・エス、ユン・ティー・ジェイ、ユー・ケイ・ワイ、ラ・ケイ・エス、キム・ジェイ・エム、他(2011)。Lactobacillus plantarum M-2 による高麗紅参の発酵とその免疫学的活性。Appl. Biochem. Biotechnol. 165, 1107-1119. doi: 10.1007/s12010-011-9328-6
PubMed Abstract|CrossRef Full Text|Google Scholar
Kwon, H. K., Jo, W. R., and Park, H. J. (2018). CY誘発免疫抑制モデルにおけるPediococcus pentosaceus(GRC-ON89A)で発酵させたC. militarisの免疫増強活性。BMC Complem. Altern. Med. 18:75. doi: 10.1186/s12906-018-2133-9
PubMed Abstract|CrossRef Full Text|Google Scholar
ラニアー、L.NK細胞認識。Annu. Rev. Immunol. 23, 225-274. doi: 10.1146/annurev.immunol.23.021704.115526
クロスレフフルテキスト|Google Scholar
Lee, N. K., and Paik, H. D. (2017). 乳酸菌を用いたバイオコンバージョン。ジンセノサイド、GABA、およびフェノール化合物。J. Microbiol. Biotechnol. 27, 869-877. doi: 10.4014/jmb.1612.12005.
PubMed Abstract|CrossRef Full Text|Google Scholar
Lee, H. J., Woo, Y., Hahn, T. W., Jung, Y. M., and Jung, Y. J. (2020).食細胞の形成と成熟。ファゴソームの形成と成熟:細胞内細菌感染に対する自然免疫の重要な機構. Microorganisms 8:1298。doi: 10.3390/microorganisms8091298
PubMed Abstract|CrossRef Full Text|Google Scholar
Levy, M., Kolodziejczyk, A. A., Thaiss, C. A., and Elinav, E. (2017). ディスバイオシスと免疫系。Nat. Rev. Immunol. 17, 219-232. doi: 10.1038/nri.2017.7
クロスレフフルテキスト|Google Scholar
Li, X. Y. (2021). 漬物からの乳酸菌の分離とスクリーニング、およびゴジベリージュースの発酵によるマウスへの免疫調節効果。[学位論文/修士論文]。[寧夏(中国)]: 寧夏大学
Google Scholar
李、W.、胡、X.、王、S.、焦、Z.、孫、T.、劉、T.、その他 (2020). ハトムギ多糖類の特性解析と免疫調節による抗腫瘍生物活性.Int. J. Biol. Macromol. 145, 985-997. doi: 10.1016/j.ijbiomac.2019.09.189.
PubMed Abstract|CrossRef Full Text|Google Scholar
Li, Y., Liu, H., Qi, H., Tang, W., Zhang, C., Liu, Z., et al. (2021). Ganoderma lucidum子実体抽出物のプロバイオティクス発酵は、デキサメタゾン誘発免疫抑制マウスにおける免疫賦活活性を促進した。Biomed. Pharmacother. 141:111909. doi: 10.1016/j.biopha.2021.111909.
PubMed Abstract|CrossRef Full Text|Google Scholar
Liang, Z. J. (2019). 発酵ハトムギ多糖類の構造的特徴およびマウスにおける腸管粘膜免疫の改善効果。[学位論文/修士論文]. [北京(中国)].中国農業科学院
Google Scholar
Liu, K. K., Wang, Q. T., Yang, S. M., Chen, J. Y., Wu, H. X., and Wei, W. (2014). ジンセノサイド化合物Kは、コラーゲン誘発関節炎マウスにおけるTリンパ球の異常な活性化を抑制する。Acta Pharmacol. Sin. 35, 599-612. doi: 10.1038/aps.2014.7
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Lodha, D., Das, S., and Hati, S. (2021). 大豆乳酸発酵中の抗酸化活性、総フェノール量およびイソフラボンの生体内変換。J. Food Process. Preserv. 45:e15583. doi: 10.1111/jfpp.15583
クロスレフフルテキスト|Google Scholar
Luo, Y., Fang, Q., Lai, Y., Lei, H., Zhang, D., Niu, H.、他 (2022). Polygonatum sibiricum redの葉からの多糖類。マウスの腸内細菌叢を制御し、短鎖脂肪酸の産生に影響を与える。AMB Express 12:35. doi: 10.1186/s13568-022-01376-z
PubMed Abstract|CrossRef Full Text|Google Scholar
Luu, M., Pautz, S., Kohl, V., Singh, R., Romero, R., Lucas, S., et al.(2019)。短鎖脂肪酸ペンタノエートは、リンパ球の代謝-エピジェネティッククロストークを調節することにより自己免疫を抑制する。Nat. Commun. 10:760. doi: 10.1038/s41467-019-08711-2
PubMed Abstract|CrossRef Full Text|Google Scholar
このような、遺伝子発現を制御するために必要な情報は、以下の通りです。免疫抑制マウスにおける機能性オリゴ糖の腸管免疫調節への影響。Saudi. J. Biol. Sci. 27, 233-241. doi: 10.1016/j.sjbs.2019.08.019.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
マクファーレン、S.、およびマクファーレン、G.T. (2003). 短鎖脂肪酸産生の制御.Proc. 62, 67-72.
クロスレフフルテキスト|Google Scholar
Mahapatro, M., Erkert, L., and Becker, C. (2021). 腸の免疫細胞と上皮細胞間のサイトカインを介したクロストーク。Cells 10:111.
PubMed Abstract|CrossRef Full Text|Google Scholar
Małaczewska, J., Kaczorek-Łukowska, E., Wójcik, R., Rękawek, W., and Siwicki, A. K. (2019). ブタの白血球に対するナイシンのin vitro免疫調節効果。J. Anim. Physiol. Anim. Nutr. 103, 882-893. doi: 10.1111/jpn.13085.
PubMed Abstract | CrossRef Full Text | Google Scholar
マレク、T. R. (2003). IL-2の主な機能は、T制御細胞の発達を促進することである。J. Leukoc. バイオロジー 74, 961-965.
PubMed Abstract|CrossRef Full Text|Google Scholar
Manach, C., Scalbert, A., Morand, C., Rémésy, C., and Jiménez, L. (2004). ポリフェノール:食物源とバイオアベイラビリティ。Am. J. Clin. Nutr. 79, 727-747.
クロスレフフルテキスト|Google Scholar
マルティンス、S.、ムサット、S.I.、マルティネス-アビラ、G.、モンタニェス-サエンス、J.、アギュラー、C.N.、およびテシェイラ、J.A.(2011年)。生物活性フェノール化合物:固相発酵による生産と抽出。A review. Biotechnol. 29, 365-373. doi: 10.1016/j.biotechadv.2011.01.008.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
マストゥバ, Y., 岡本, K., 加藤, Y., 山口, N., そして赤澤-工藤, S. (2017). 乳酸菌が設計した発酵民族薬(プエラリア・ミリフィカ)が補体成分の活性化を介して白血球サブセットを制御した。Open J. Rheumatol. Autoimm. Dis. 07, 65-79. doi: 10.4236/ojra.2017.71006
クロスレフフルテキスト|Google Scholar
Min, L., Wang, H., and Qi, H. (2022). アストラガロシドIVは、TLR4/NF-κB/STAT3シグナル経路を介してマクロファージ分極を調節することにより、肝癌の進行を抑制する。Am. J. Transl. Res. 14, 1551-1566.
Google Scholar
Mkaddem, S. B., Christou, I., Rossato, E., Berthelot, L., Lehuen, A., and Monteiro, R. C. (2014). IgA、IgA受容体、およびそれらの抗炎症性。Curr.Top. Microbiol. Immunol. 382, 221-235.
PubMed Abstract|CrossRef Full Text|Google Scholar
Mo, S. J., Nam, B., Bae, C. H., Park, S. D., Shim, J. J., and Lee, J. L. (2021).新規乳酸菌の特性評価。紅参抽出物を用いた生理特性改善および免疫増強効果のある新規乳酸菌パラカゼイHY7017の性状解析。発酵 7:238. doi: 10.3390/fermentation7040238
クロスレフフルテキスト|Google Scholar
Moein, M., Imani Fooladi, A. A., and Mahmoodzadeh Hosseini, H. (2018). マクロファージ細胞に対するグリーンケミストリーで合成されたアグ-ナイシンナノパーティクルの効果の判定。Microb. Pathog. 114, 414-419. doi: 10.1016/j.micpath.2017.12.034.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
村上 浩、大河原 浩、廣瀬 浩(2004).多発性骨髄腫患者におけるTh1/Th2細胞.血液学 9, 41-45.
PubMed Abstract|CrossRef Full Text|Google Scholar
Murphy, E. J., Rezoagli, E., Major, I., Rowan, N. J., and Laffey, J. G. (2020).を参照。β-グルカンの代謝・免疫調節特性と臨床応用の可能性. J. Fungi 6:356. doi: 10.3390/jof6040356
PubMed Abstract|CrossRef Full Text|Google Scholar
Nguyen, M. T., and Nguyen, T. M. T. (2015). 霊芝からの生理活性化合物の抽出と胞子粉の採取。CTUJS 01, 53-60. doi: 10.22144/ctu.jen.2015.001.
クロスレフフルテキスト|Google Scholar
Nwodo, U. U., Green, E., and Okoh, A. I. (2012). 細菌性エキソ多糖:機能性と展望。Int. J. Mol. Sci. 13, 14002-14015.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
オッフェルマンズ、S. (2017). 代謝におけるヒドロキシカルボン酸受容体の作用. Trends Endocrinol. Metab. 28, 227-236. doi: 10.1016/j.tem.2016.11.007.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Özcelik, S., Kuley, E., and Özogul, F. (2016). 乳酸菌による乳酸、酢酸、コハク酸、プロピオン酸、ギ酸および酪酸の生成。LWT 73, 536-542. doi: 10.1016/j.lwt.2016.06.066.
クロスレフフルテキスト|Google Scholar
Pang, Z., Chen, J., Wang, T., Gao, C., Li, Z., Guo, L., et al. (2021). 植物二次代謝産物と植物マイクロバイオームのリンク:レビュー。Front. Recent Dev. Plant Sci. 12:621276.
PubMed Abstract|CrossRef Full Text|Google Scholar
朴、C. M., 金、G. M., および茶、G. S. (2021). キムチから新たに分離し、特徴付けた乳酸菌ペントサスNGI01株によるフラボノイドのバイオトランスフォーメーション。Microorganisms 9:1075。doi: 10.3390/microorganisms9051075
PubMed Abstract|CrossRef Full Text|Google Scholar
Park, S. Y., Kim, H. B., Kim, J. H., Lee, J. M., Kim, S. R., Shin, H. S., and al. (2014). マウスモデルにおける発酵紅参の免疫賦活効果。Prev. Nutr. Food Sci. 19, 10-18. doi: 10.3746/pnf.2014.19.1.010.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
パテル、S.およびゴヤル、A. (2011). 機能性オリゴ糖:生産、特性および応用。を参照。Biotechnol. 27, 1119-1128. doi: 10.1007/s11274-010-0558-5
クロスレフフルテキスト|Google Scholar
Peters, A., Krumbholz, P., Jäger, E., Heintz-Buschart, A., Çakir, M. V., Rothemund, S., et al. (2019). 発酵食品に存在する乳酸菌の代謝産物は、ヒトヒドロキシカルボン酸受容体3の非常に強力なアゴニストである。PLoS Genet. 15:e1008145. doi: 10.1371/journal.pgen.1008145
PubMed Abstract|CrossRef Full Text|Google Scholar
Peters, V., van de Steeg, E., van Bilsen, J., and Meijerink, M. (2019). プレおよびプロバイオティクスのメカニズムと免疫調節特性. ベネフィック. Microbes 10, 225-236. doi: 10.3920/BM2018.0066.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Prud'homme, G. J., Glinka, Y., and Wang, Q. (2015). 自己免疫疾患における免疫学的なGABA作動性相互作用と治療への応用. Autoimmun. Rev. 14, 1048-1056. doi: 10.1016/j.autrev.2015.07.011
クロスレフフルテキスト|Google Scholar
Rajoka, M. S. R., Wu, Y., Mehwish, H. M., Bansal, M., and Zhao, L. (2020). Lactobacillus exopolysaccharides: engineering strategies, physiochemical functions, and immunomodulatory effects on host healthに関する新しい視点. Trends Food Sci. Technol. 103, 36-48. doi: 10.1016/j.tifs.2020.06.003
クロスレフフルテキスト|Google Scholar
Ren, C., Dokter-Fokkens, J., Figueroa Lozano, S., Zhang, Q., de Haan, B. J., Zhang, H., et al(2018)。乳酸菌は杯細胞の調節により腸管バリア機能に影響を与える可能性がある。モル. Nutr. Food Res. 62:e1700572. doi: 10.1002/mnfr.201700572.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Ricci, A., Cirlini, M., Calani, L., Bernini, V., Neviani, E., Del Rio, D., et al.(2019)。乳酸菌によるエルダーベリージュースポリフェノールのin vitro代謝(In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria). Food Chem. 276, 692-699. doi: 10.1016/j.foodchem.2018.10.046.
PubMed Abstract|CrossRef Full Text|Google Scholar
Rochman, Y., Spolski, R., and Leonard, W. J. (2009). γ(c)ファミリーサイトカインによるT細胞の制御に関する新たな洞察。Nat. Rev. Immunol. 9, 480-490.
PubMed Abstract | CrossRef Full Text | Google Scholar
このような状況下において、「大腸菌の代謝産物」と「ヒトの健康」はどのように関わっているのだろうか。大腸菌の代謝産物とヒトの健康。Curr. Opin. Microbiol. 16, 246-254. doi: 10.1016/j.mib.2013.07.002
クロスレフフルテキスト|Google Scholar
Schenten, D., and Medzhitov, R. (2011). 自然免疫系による適応免疫応答の制御。Adv. Immunol. 109, 87-124. doi: 10.1016/B978-0-12-387664-5.00003-0
クロスレフフルテキスト|Google Scholar
Song, M. W., Park, J. Y., Lee, H. S., Kim, K. T., and Paik, H. D. (2021).乳酸菌による共培養。ラクトバチルス・ブレビスB7による共発酵は、水耕人参強化ヨーグルトの抗酸化および免疫調節活性を向上させる。アンチオキシダント10:1447.doi: 10.3390/antiox10091447
PubMed Abstract|CrossRef Full Text|Google Scholar
Usta-Gorgun, B., and Yilmaz-Ersan, L. (2020). ビフィドバクテリウム属菌によるサレップ存在下での短鎖脂肪酸の生産。Electron. J. Biotechnol. 47, 29-35. doi: 10.1016/j.ejbt.2020.06.004.
クロスレフフルテキスト|Google Scholar
Vivier, E., Tomasello, E., Baratin, M., Walzer, T., and Ugolini, S. (2008). ナチュラルキラー細胞の機能。Nat. Immunol. 9, 503-510.
クロスレフフルテキスト|Google Scholar
Wang, J., Fang, X., Wu, T., Fang, L., Liu, C., and Min, W. (2020). 乳酸菌プラネタリウムJLAU103が生産する酸性エクソポリサッカライドのRAW264.7マクロファージに対するin vitro免疫調節効果(In vitro immunomodulatory effects of acidic exopolysaccharide produced by lactobacillus planetarium JLAU103 on RAW264.7 macrophages. Int. J. Biol. Macromol. 156, 1308-1315. doi: 10.1016/j.ijbiomac.2019.11.169
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, J., Feng, W., Tang, F., Ao, H., and Peng, C. (2019). 生薬から分離された4つのグループの天然化合物の治療活性の潜在的な改善要因である腸内細菌の形質転換。フィトテラピー 138:104293. doi: 10.1016/j.fitote.2019.104293.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Wang, J., and Fu, Y. X. (2004). T細胞媒介性免疫におけるLIGHTの役割。Immunol. 30, 201-214.
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, T., and He, C. (2020). TNF-αとIL-6:免疫系と骨系の関連性。Curr. Drug Targets 21, 213-227. doi: 10.2174/1389450120666190821161259
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, S., Huang, S., Ye, Q., Zeng, X., Yu, H., Qi, D., et al. (2018). 抗菌ペプチドSublancinによるシクロホスファミド誘発性免疫抑制のマウスへの予防。J Immunol Res 2018:4353580, -4353511. doi: 10.1155/2018/4353580
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, Y., Qin, Q., Chen, J., Kuang, X., Xia, J., Xie, B.、他(2009). また、このような臓器移植は、臓器移植を行う臓器がない場合にも行うことができる。Transplant. Immunol. 22, 5-11. doi: 10.1016/j.trim.2009.09.004
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, M., Yan, S. J., Zhang, H. T., Li, N., Liu, T., Zhang, Y. L., et al.(2017)。Ginsenoside Rh2はメラノーママウスモデルの抗腫瘍免疫反応を増強する。オンコル.Lett. 13, 681-685. doi: 10.3892/ol.2016.5490.
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, S., Ye, Q., Wang, K., Zeng, X., Huang, S., Yu, H., et al.(2019)。抗菌ペプチドSublancinによるマクロファージ機能の強化は、メチシリン耐性黄色ブドウ球菌からマウスを保護する。J. Immunol. Res. 2019:3979352. doi: 10.1155/2019/3979352.
PubMed Abstract|CrossRef Full Text|Google Scholar
Wang, H., Yin, H., Zhong, Y., Hu, J., Xia, S., Wang, Z., et al.(2022)。発酵コイキング種子由来の多糖類は、腸内細菌叢を変化させることにより、循環窒素と免疫機能を調節する。Curr. Res. 5, 1994-2003. doi: 10.1016/j.crfs.2022.10.007.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
渡辺義明・山口直樹・堀内郁夫・村山毅(2015). 乳酸菌が設計した発酵黒ウコンが白血球サブセットと抗酸化活性をリアレンジした。Open J. Immunol. 05, 199-214. doi: 10.4236/oji.2015.53016.
クロスレフフルテキスト|Google Scholar
Wen, A., Xie, C., Mazhar, M., Wang, C., Zeng, H., Qin, L., et al.(2020)。Bacillus subtilisによるハトムギ(Coix lacryma-jobi)由来のTetramethylpyrazineの生変換とその品質特性. J. Food Sci. Technol. 57, 4092-4102. doi: 10.1007/s13197-020-04443-0
PubMed Abstract|CrossRef Full Text|Google Scholar
ウィリアムソン、G. (2017). 現代栄養学におけるポリフェノールの役割。Nutr.ブル. 42, 226-235. doi: 10.1111/nbu.12278.
PubMed Abstract|CrossRef Full Text|Google Scholar
楊、M.、孟、F.、顧、W.、福、L.、張、F.、李、F.、他(2021)。Polygonatum kingianum由来の多糖類がラクトバチルス・フェイシスの短鎖脂肪酸産生およびクオラムセンシングに与える影響。Front. Microbiol. 12:758870。doi: 10.3389/fmicb.2021.758870
PubMed Abstract|CrossRef Full Text|Google Scholar
Yang, W., Yu, T., Huang, X., Bilotta, A. J., Xu, L., Lu, Y., et al.(2020)。腸内細菌叢由来の短鎖脂肪酸による免疫細胞IL-22産生と腸管免疫の制御。Nat. Commun. 11:4457. doi: 10.1038/s41467-020-18262-6
PubMed Abstract|CrossRef Full Text|Google Scholar
Yin, Z., Liang, Z., Li, C., Wang, J., Ma, C., and Kang, W. (2021). 食用キノコ由来の多糖類の免疫調節効果:レビュー。Food Sci. Hum. Wellness 10, 393-400. doi: 10.1016/j.fshw.2021.04.001.
クロスレフフルテキスト|Google Scholar
Yin, M., Zhang, Y., and Li, H. (2019). 植物多糖類によるマクロファージの免疫制御に関する研究の進展. Front. Immunol. 10:145. doi: 10.3389/fimmu.2019.00145
PubMed Abstract|CrossRef Full Text|Google Scholar
Yoo, J., Lee, J., Zhang, M., Mun, D., Kang, M., Yun, B., et al(2022)。発酵鹿角ビロードのγ-アミノ酪酸とシアル酸の増強と免疫促進効果. J. Anim. Sci. Technol. 64, 166-182. doi: 10.5187/jast.2021.e132
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Zacharof, M. P., and Lovitt, R. W. (2012). 乳酸菌が産生するバクテリオシンの総説。Apcbee Proc. 2, 50-56.
クロスレフフルテキスト|Google Scholar
このように、樹状細胞は、その増殖や分化の過程において、様々な役割を担っています。樹状細胞のレビュー、臨床免疫学における役割、様々な動物種における分布。IJMS 22:8044.論文番号: 10.3390/ijms22158044
PubMed Abstract|CrossRef Full Text|Google Scholar
この論文では、フルクタンとガラクトシルグリセロールの構造について解説しています。また、このような環境下において、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」を実現するためには、「環境負荷の低減」が不可欠である。Carbohydr. Polym. 267:118219.
PubMed Abstract|CrossRef Full Text|Google Scholar
Zhang, C., He, J., Wang, X., Su, R., Huang, Q., Qiao, F., et al.(2022)。食餌性γ-アミノ酪酸(GABA)は、中国のミテガニ(Eriocheir sinensis)稚魚の非特異的免疫を改善し、リポポリサッカライド(LPS)誘発免疫過剰反応を緩和させる。魚貝類免疫学. 124, 480-489.
PubMed Abstract|CrossRef Full Text|Google Scholar
このような背景のもとで、私たちは、「食の安全」、「食の安心」、「食の豊かさ」、「食の楽しさ」、「食の豊かさ」を追求し、「食の豊かさ」を実現するための研究を進めています。腸内における機能性オリゴ糖発酵:腸の健康増進とその決定要因-レビュー。Carbohydr. Polym. 284:119043. doi: 10.1016/j.carbpol.2021.119043
PubMed Abstract|CrossRef Full Text|Google Scholar
このように、ノニ醗酵液は、その性質上、醗酵の進行に伴い、醗酵が進行する。ノニ滲出液で処理した樹状細胞は、Bリンパ球の増殖と分化を直接的に刺激する。Oncol. Rep. 21, 1147-1152. doi: 10.3892/or_00000334
クロスレフフルテキスト|Google Scholar
Zhao, Y., Hong, K., Zhao, J., Zhang, H., Zhai, Q., and Chen, W. (2019). Lactobacillus fermentumとその潜在的な免疫調節特性。J. Funct. Foods 56, 21-32. doi: 10.1016/j.jff.2019.02.044.
クロスレフフルテキスト|Google Scholar
Zhao, Y., Zhong, X., Yan, J., Sun, C., Zhao, X., and Wang, X. (2022). 天然物の生体内変換における腸内細菌の潜在的役割:概説。Front. Microbiol. 13:956378.
PubMed Abstract|CrossRef Full Text|Google Scholar
Zhou, X., and Ye, Q. (2021). COVID-19に対する細胞性免疫応答と潜在的な免疫モジュレーター。Front. Immunol. 12:646333. doi: 10.3389/fimmu.2021.646333.
PubMed Abstract|CrossRef Full Text|Google Scholar
Zhu, F. J., Tong, Y. L., Sheng, Z. Y., and Yao, Y. M. (2019). バイオマテリアルに対する宿主応答における樹状細胞の役割とそのシグナル伝達経路. Acta Biomater. 94, 132-144. doi: 10.1016/j.actbio.2019.05.038.
PubMed Abstract|CrossRefフルテキスト|Google Scholar
Zong, Q. H., Lou, Y. L., Zhang, H., and Wang, S. S. (2018). 脳損傷マウスにおけるプロバイオティクスFGMで発酵させたハトムギ多糖の免疫学的効果。Chin. J. Immunol. 34, 36-39. doi: 10.3969/j.issn.1000-484X.
クロスレフフルテキスト|Google Scholar
Zwirner, N. W., Domaica, C. I., and Fuertes, M. B. (2021). 感染症および癌におけるNK細胞の制御機能。J. Leukoc. Biol. 109, 185-194. doi: 10.1002/JLB.3MR0820-685R
PubMed Abstract|CrossRef Full Text|Google Scholar
キーワード:乳酸菌、生薬、発酵、免疫調節、生理活性物質、メカニズム
引用元 Zhu H, Guo L, Yu D and Du X (2022) New insights into immunomodulatory properties of lactic acid bacteria fermented herbal medicines. Front. Microbiol. 13:1073922.doi: 10.3389/fmicb.2022.1073922
Received: 2022年10月19日; Accepted: 2022年11月10日
公開:2022年11月28日
編集者
ローラン・デュフォッセ、レユニオン大学、フランス
査読者
Xin Cao, Jilin Agricultural University, China(中国
Xiao Wang, Inner Mongolia University, China(内モンゴル大学、中国
Copyright © 2022 Zhu, Guo, Yu and Du. これは、クリエイティブ・コモンズ表示ライセンス(CC BY)の条件の下で配布されるオープンアクセス論文である。原著者および著作権者のクレジットを表示し、本誌の原著を引用することを条件に、他のフォーラムでの使用、配布、複製を許可する。本規定に従わない使用・配布・複製は認めない。
*Correspondence: Xiaowei Du, xiaoweidu@hotmail.com
免責事項:本論文で表明されたすべての主張は,あくまでも著者のものであり,必ずしも所属機関のもの,あるいは出版社,編集者,査読者のものを代表するものではありません。この記事で評価される可能性のある製品、またはそのメーカーが行う可能性のある主張は、出版社によって保証または承認されるものではありません。
こんな人も見ています
Lactobacillus paracasei,Lactobacillus plantarum,Lactobacillus brevisで発酵させたKamut®,キヌアおよび小麦の生地における香料の生成。SPME-GC/MSによる研究
Tiziana Di Renzo, Anna Reale, Floriana Boscaino, Maria C. Messia(ティツィアナ ディ レンゾ、アンナ レアーレ、フロリアナ ボスキーノ、マリア C. メシア
エスニック発酵乳製品に含まれる乳酸菌のプロバイオティクス特性に関する機構的知見
Tamoghna Ghosh, Arun Beniwal, Anupama Semwal, Naveen Kumar Navani
Lacticaseibacillus rhamnosus ZFM216株が産生するバクテリオシンの精製と性状解析
Danli Wu, Mengdi Dai, Yongqing Shi, Qingqing Zhou, Ping Li, Qing Gu(呉丹莉、戴萌迪、周慶慶、李平、顧慶
シクロフォスファミド誘発マウスにおけるAuricularia cornea var. Li.多糖類の免疫系および腸内細菌叢に対する制御作用
趙明、石偉、陳西軍、劉延芳、楊燕、孔祥慧
Kanamycin Bに応答するトランスクリプトームプロファイリングにより、Escherichia coliにおけるその幅広い非抗生細胞機能が明らかになった。
Yaowen Chang, Xuhui Zhang, Alastair I. H. Murchie, Dongrong Chen(張耀文、張旭輝、Alastair I. H. Murchie、陳東栄
この記事が気に入ったらサポートをしてみませんか?