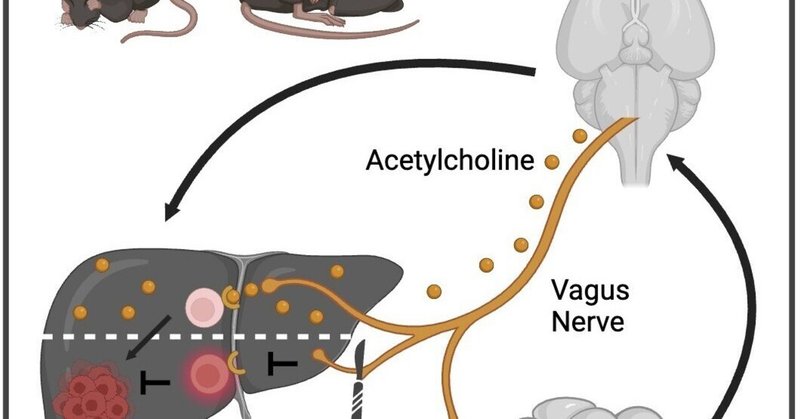
腸内マイクロバイオームは迷走神経を介して肝腫瘍を制御する

本文へスキップ
バイオレクシブ
ホーム投稿FAQブログアラート / RSSチャンネルについて
このキーワードで検索
検索
詳細検索
新着結果 このプレプリントをフォローする
腸内マイクロバイオームは迷走神経を介して肝腫瘍を制御する
https://www.biorxiv.org/content/10.1101/2024.01.23.576951v1.full

View ORCID ProfileKylynda C. Bauer, View ORCID ProfileRajiv Trehan, View ORCID ProfileBenjamin Ruf, View ORCID ProfileYuta Myojin, View ORCID ProfileMohamed-Reda Benmebarek, Chi Ma, View ORCID ProfileMatthias Seifert, Amran Nur, Jonathan Qi, Patrick Huang, Marlaine Soliman, Benjamin L. Green, View ORCID ProfileSimon Wabitsch, Danielle A. Springer, Francisco J. Rodriguez-Matos, Shadin Ghabra, View ORCID ProfileStephanie N. Gregory, Jennifer Matta, Brian Dawson, View ORCID ProfileJihye Golino, Changqing Xie, View ORCID ProfileAmiran Dzutsev, View ORCID ProfileGiorgio Trinchieri, View ORCID ProfileFirouzeh Korangy, View ORCID ProfileTim F. Greten
doi: https://doi.org/10.1101/2024.01.23.576951
この論文はプレプリントであり、査読による認証を受けていない[これはどういう意味か?]
00000019
要旨全文情報/履歴メトリクスプレビューPDF
要旨
肝臓がんは最も死亡率の高いがんのひとつである。神経は、腫瘍進行の制御因子として控えめに研究されてきた。副交感神経である迷走神経は、アセチルコリン(ACh)を介して全身の免疫に影響を及ぼす。コリン作動性の神経免疫相互作用が肝細胞癌(HCC)に影響を及ぼすかどうかはまだ不明である。肝迷走神経切断術(HV)による肝神経脱神経は肝腫瘍負荷を有意に減少させたが、一方で副交感神経緊張の薬理学的増強は腫瘍増殖を促進した。Rag1KOマウスにおけるコリン作動性破壊により、コリン作動性調節には適応免疫が必要であることが明らかになった。さらに、scRNA-seqとin vitro研究により、迷走神経AChがムスカリンACh受容体(AChR)CHRM3を介してCD8+ T細胞の活性を減弱させることが示された。 CD8+ T細胞の枯渇はHVの結果を無効にし、CD8+ T細胞上のChrm3の選択的欠失は肝腫瘍増殖を抑制した。腫瘍特異的な転帰にとどまらず、迷走神経切断術はがんに伴う疲労や不安様行動を改善した。肝細胞癌ドナーからの微生物叢移植は行動を障害するのに十分であったので、我々は微生物叢と神経免疫の推定的クロストークを調べた。迷走神経切断よりもむしろ腫瘍の方が、糞便中の細菌組成を強固に変化させ、DesulfovibrionalesとClostridial分類群を増加させた。驚くべきことに、腫瘍のないマウスでは、迷走神経切断によって肝細胞癌に関連した微生物叢が肝CD8+T細胞を活性化することが認められた。これらの知見は、腸内細菌が、ダイナミックで薬剤学的に標的化可能な迷走神経-肝臓軸を介して、行動と肝臓の抗腫瘍免疫に影響を及ぼすことを明らかにした。
はじめに
原発性肝がんは、依然として世界的ながん関連死の主要原因である(1, 2)。さらに、肝臓は腫瘍転移の最も多い臓器の一つである(3)。肝免疫抑制は癌の予後不良の一因であり、様々な癌種において死亡率を悪化させる(4)。肝臓は著しく免疫寛容な臓器である(5)。腸由来の抗原にさらされ続ける肝臓は、積極的に炎症を抑制する(5, 6)。このような環境は腫瘍の急速な増殖を促進し、免疫療法の効果を損なう(7-9)。
免疫浸潤以外にも、末梢の神経が腫瘍の微小環境を支配し、形成している(10, 11)。直接的な神経-腫瘍シグナル(12-14)や間接的な神経免疫回路(15)が腫瘍負荷に影響を及ぼしている可能性が高い。がん神経科学という新たな概念は、このようなダイナミックで双方向性の、しかもほとんど研究されていない相互作用を捉えている(16)。ここでは、迷走神経活動(迷走神経➔肝、肝➔脳)の双方向性が肝腫瘍の転帰をどのように制御するかを評価する。
迷走神経(脳神経X)は副交感神経系の主要な構成要素であり、ACh活性を介して内臓器官を調節する(17、18)。コリン作動性迷走神経活動は、乳癌(19)、大腸癌(20)、胃癌(21)、膵癌(22)、前立腺癌の進行に影響を及ぼす(23)。最近では、迷走神経支配が肝癌の予後と病理に関与している(24-26)。コリン作動性神経密度は肝細胞癌患者の予後不良と相関しており(26)、消化性潰瘍の治療を受けた約50,000人の患者を対象としたレトロスペクティブ研究では、縫合術(非脱神経)と比較して、三半規管迷走神経切断術(腹部脱神経)後に肝癌や胆道癌のリスクが低下することが報告されている(24)。マウスHCCモデルでは、非迷走神経コリン合成がCD4+ T細胞の免疫監視と消耗を制御していた。CD4+ T細胞におけるコリンアセチルトランスフェラーゼ(Chat)の遺伝子異常は肝腫瘍負荷を悪化させた(25)。対照的に、コリン作動性迷走神経活動は、マクロファージ(敗血症モデル)(27)および腫瘍関連マクロファージ(膵臓がんモデル)(22)による炎症性サイトカインの放出を減少させた。この抗炎症性調節が腫瘍微小環境を形成している可能性が高い(27)。このコリン作動性抗炎症アークが肝抗腫瘍免疫に影響するかどうか、またどの程度影響するかはまだ不明である。
我々は、マウスの外科的肝枝迷走神経切断術が肝腫瘍負荷を有意に減少させたことを報告する。迷走神経切断された肝臓は広範な炎症反応を示し、コリン作動性抗炎症アークを支持した(27, 28)。転写解析と軌跡解析により、CD8+T細胞サブセット間で異なるAch制御が明らかになった。我々は、AChシグナルがCD8+ T細胞のエフェクター機能に影響を与えることを立証した。実際、コリン作動性活性はCD8+ T細胞の炎症、細胞傷害性、腫瘍媒介性殺傷に影響を及ぼし、一方、CD8+ T細胞上のAChR Chrm3の遺伝子欠損は肝細胞癌の増殖を抑制した。偽手術対照と比較して、HVマウスはまた、微生物異常症と関連する行動であるがん性疲労と不安様反応の減少を示した。肝細胞癌微生物叢移植により、腸内細菌が迷走神経を介して肝抗腫瘍免疫を制御していることが明らかになった。この腸-脳アークは、神経免疫誘導療法および肝がん治療の潜在的標的を同定するものである。
研究結果
肝迷走神経切断はマウスの肝腫瘍負担を軽減する
肝癌における迷走神経の影響を調べるために、マウスは外科的肝迷走神経切断術(HV)または偽手術(SV)を受けた(29)。総肝迷走神経枝の切断は、目視およびH&E組織学的に確認した(図1A)。肝迷走神経切断後、体重および肝重量、ならびに肝組織像に変化は認められなかった(図s1A-C)。
図S1
図をダウンロード
新しいタブで開く
図S1
肝迷走神経切断は、代謝の変化とは無関係に、複数のマウスモデルにおいて肝腫瘍に影響を及ぼす
(A)非腫瘍の偽マウス(SVx)と肝迷走神経切断マウス(HVx)の体重。(B)SVxおよびHVxマウスの肝臓重量。(C)代表的なSVxおよびHVxマウスの肝臓の肉眼組織像およびH&E組織像。(D)非腫瘍マウス(n=SVx 7匹、HVx 7匹)の血清からの肝機能測定。灰色のオーバーレイは基準範囲を示す。(E)RIL175のHVsおよびSVs肝組織からのVEGFaサイトカインレベル、データは組織重量で正規化(n = 7 SVx, 7 HVx)。C-EのマウスはAおよびBで報告されたコホートのもの。(F)(左-右)RIL175モデル(n=20 SV、19 HV)におけるマウスの体重、腫瘍のない肝臓重量、および腫瘍:体比。(G)腫瘍発生21日後の雄RIL175マウスにおける腫瘍重量(n=11 SV、15 HV)。(H)(左-右)B16-F10モデルにおけるマウスの体重、腫瘍のない肝臓重量、および腫瘍:体比(n=16 SV、15 HV)。(I)(左-右)マウスの体重、A20モデルの表面転移腫瘍(MET)の総数、最も広い領域で測定した最大のA20 METの直径、およびA20腫瘍肝臓:体重。METsの測定は盲検化された研究者によって決定された(n = 15 SV, 15 HV)。(I) A、BおよびF-Iのデータは、2回の独立したプール実験から成る。棒グラフは平均値±SEM。統計的有意性は対にしないスチューデントのt検定によって決定した: *P < 0.05; **P < 0.01; ***P < 0.005; ns = 有意でない。
図1
図をダウンロード
新しいタブで開く
図1
特異的肝枝迷走神経切断術は肝癌の腫瘍負担を軽減する
(A)特異的肝枝迷走神経切断術と神経切断のH&E組織図。
(B)腫瘍癌モデル: C57BL/6マウスは、偽手術(SVマウス)または肝枝迷走神経切断術(HVマウス)の後、RIL175(肝細胞癌)またはB16-F10(メラノーマ)腫瘍細胞の肝内注入を受けた。また、BALB/cマウスの尾静脈からA20(リンパ腫)細胞を注射した。文字の色は腫瘍モデルを表す。特に断りのない限り、腫瘍負荷と肝免疫組成は腫瘍発生から21日後に評価した。(C)ルシフェラーゼ発現RIL175腫瘍の代表的なin vivoイメージング(IVIS® Spectrum)。腫瘍の成長に伴う発光(n = SV 10, HV 9)。(D)RIL175モデルの腫瘍重量と代表画像。データはCで示したマウスを含み、n = SV 20匹、HV 19匹。(E)B16-F10モデルの腫瘍重量と代表画像(n = 16 SV, 15 HV)。(F)A20モデル(n=15 SV、15 HV)の転移腫瘍(METs)径と代表画像。白星は腫瘍結節を示す。(G)RIL175脇腹皮下モデルの腫瘍重量と代表画像(n = 18 SV, 18 HV)。D-Gのデータは2回の独立した実験の平均±SEMを表す。(H) Co-detection by indexing (CODEX) multiplexed immunofluorescent imagingを、(36)で我々のグループにより報告された12例の肝細胞癌(HCC)患者の外科的切除に対して行った。LHCC35生検からの、平滑筋(αSMA+)、内皮細胞(CD34+)、胆管(panCK+、EpCAM+、NCAM+)、免疫(CD45+)細胞のマーカーを伴う肝神経(NCAM+)を示す代表的な6色画像。(I)HCC腫瘍隣接検体におけるHCC手術検体12検体にわたる最近傍プロファイル。最近傍の頻度:18色の抗体パネル(補足ファイル2参照)を用いて、神経に沿った3箇所で決定された最近傍の細胞のある同じ大きさの領域から行われた手動の同定解析。上の長方形は、細胞タイプ間の最近接の相対的平均距離(μm)を示す。下の円は神経/生検領域の相対的な量を示す。統計的有意性は対にしないスチューデントのt検定で決定した: *P < 0.05; **P < 0.01; ns = 有意ではない。αSMA = α-平滑筋アクチン、DC = 樹状細胞、EpCAM = 上皮細胞接着分子、LYVE1 = リンパ管内皮ヒアルロン酸受容体1、NCAM = 神経細胞接着分子、panCK = 汎サイトケラチン、TREG = 制御性T細胞、UITC = 非従来型および自然免疫様T細胞。
迷走神経活性は全身の代謝過程と血管新生に関与しているが(10, 30)、迷走神経切断マウスはトランスアミナーゼとアルブミン肝機能を測定する血清化学検査で異常を示さなかった(図s1D)。肝血管内皮増殖因子A(VEGFa)の血清レベルも迷走神経切断マウスと偽コントロールで同等であった(図s1E)。
HVマウスは複数の肝腫瘍モデルで有意に腫瘍が小さかった(図1B-D)。雌性C57BL/6 HVマウスでは、同種RIL175(HCC)細胞の肝内注入後、腫瘍の成長と負荷の減少が観察された(図1C、D、および図s1F)。肝がんの発症には性差があることが報告されているので(31)、雄マウスでの所見を繰り返し確認した(図s1G)。我々はまた、確立された転移モデルにおいてHVの腫瘍縮小を観察した(32-34)。SVコントロールと比較して、HV肝臓はC57BL/6マウス内でより小さなB16-F10(メラノーマ)腫瘍を発症し(図1Eおよび図s1H)、BALB/c動物は尾静脈注射後、より少なく小さなA20(リンパ腫)転移腫瘍を示した(図1Fおよび図s1I)。さらに、皮下への腫瘍注入も行った。肝迷走神経切断術は皮下のRIL175腫瘍増殖に影響を与えず、肝臓特異的な結果を強調した(図1G)。まとめると、肝迷走神経切断術は、細胞株、腫瘍モデル、マウスの性別/系統に関係なく、肝腫瘍をしっかりと制御した。
肝腫瘍内の腫瘍微小環境における神経活動の因果的役割を探るために、われわれは、公開されているデータセット(TCGA [The Cancer Genome Atlas] およびGTex [Genotype-Tissue Expression] (35))で、8つの異なるヒトがん組織から得られた差次発現遺伝子(DEG)を解析した。パスウェイ濃縮解析の結果、肝臓(HCC)を含む8つのがん組織のうち7つのがん組織で神経経路が有意に変化していることが明らかになり、リム(腫瘍隣接)組織内での神経炎症と神経突起の増加が確認された(図s2A、Supplemental File 1)。また、最近われわれの研究室によって特徴づけられた小規模な臨床コホート(図1Hおよび図s2B)においても、肝細胞癌リム内の神経浸潤が観察された(36)。CODEX(索引付けによる共検出)を用いた空間免疫アトラスにより、12のHCC外科切除例でNCAM+(神経細胞接着分子)神経が検出された。神経(50μm以上の枝、DAPI-NCAM+)近傍を手作業でプロファイリングした(図1I)。38マーカーCODEXパネル((36)参照)では正確な神経細胞の同定はできないが、特異的な免疫、間質、胆道の特徴を同定した(図s2Cおよび補足ファイル2)。適応免疫細胞(CD45+CD3+)は、HCC手術標本全体で平均すると、最近接細胞の約42%を占め、最大の割合を占めた(図1Iおよび図s2D)。末梢の神経が適応免疫活性を制御していることから(15, 25)、次にコリン作動性の神経免疫相互作用がHVの転帰を形成しているかどうかを評価した。
図S2
図をダウンロード
新しいタブで開く
図S2
がん集団全体に存在する神経免疫相互作用
(A)TCGAとGTexのデータセットから得られたDEG解析から解析された、健常組織と腫瘍隣接検体から得られた7つの臨床がんにわたるパスウェイ濃縮解析。Padj値でランク付けされた、解析から得られた神経GOパスウェイのトップ。癌の種類は、( )内に差次的に発現したパスウェイの総数とともに上部に記載されている。(B)拡大されていないCODEXグラフの例、白四角はその後の拡大(100倍、図S2C参照)を示す。(C)Bで報告した生検からの代表的な構造(HNF4a、LYVE1、NCAM、汎サイトケラチン、αSMA、CD34、CD45)、リンパ球(NCAM、CD45、CD3、CD19、CD4、FoxP3、CD25、CD161、TCRVα)、骨髄球(NCAM、CD45、CD11b、CD68、CD11c)染色。細胞核はDAPI染色により同定した。(D)CODEXイメージングにより決定された12個のHCC生検サンプルの平均最近傍。CODEXデータセットと抗体クローンは(36)に記載。αSMA = α-平滑筋アクチン、EpCAM = 上皮細胞接着分子、GTex = Genotype-Tissue Expression、LYVE1 = リンパ管内皮ヒアルロン酸受容体1、NCAM = 神経細胞接着分子、 panCK = pan-Cytokeratin、TCGA = The Cancer Genome Atlas、TREG = regulatory T cell、UITC = unconventional and innate-like T cell(MAIT = mucosal-associated invariant T cellを含む)。
ACh活性はCD8+T細胞を介して肝腫瘍負荷を制御する
副交感神経迷走神経は、非肝腫瘍モデルにおいて、ACh放出を介した腫瘍増殖の阻止または促進という多様な結果を示す(19-23)。予想されたように、肝細胞癌の肝臓は偽コントローラに比べてAChレベルが低下していた(図2A)。コリン作動性作用を特異的に確認するため、膵臓がんモデルで腫瘍細胞増殖を変化させた(22)広範なムスカリン作動性AChRアゴニストであるベタネコール(図2B)でマウスを処理した。
図2
図をダウンロード
新しいタブで開く
図2
肝迷走神経切断術はコリン作動性シグナルとCD8+ T細胞を介して肝腫瘍を制御する
(A)ELISAで測定した肝AChレベルを組織タンパク質含量で正規化した。データはRIL175腫瘍発生から21日後に評価した。(B)全身性ベタネコール投与モデル:ベタネコール(400μg/ml飲料水)を腫瘍発生3-5日前に投与し、RIL175肝内注射後21日目に腫瘍負荷を評価した。(C)C57BL/6(WT)およびRag1KO(RKO)ベハネコール(BETH)モデルの腫瘍重量と代表的な肝臓画像。2つの独立したコホートからプールしたデータ、n = 16 B6_H2O, 11 B6_BETH, 13 RKO_H2O, 13 RKO_BETH。(D) フローサイトメトリーで評価した肝免疫細胞組成、平均頻度(生細胞%)、n = 10 SV, 9 HV。(E)組織重量で正規化したDからの肝CD8+ T細胞の細胞数。(F)CD8+T細胞抗体枯渇腫瘍モデル: C57BL/7マウスを無作為に割り付け、IgG2bコントロールまたは抗CD8抗体(200μg/マウス、肝内注射の1日前から週1回)を投与した。(G)21日後のRIL175腫瘍重量と代表的な肝臓画像、n=7 SV-IgG、8 HV-IgG、9 SV-aCD8、8 HV-aCD8。A、C、G、Eのデータは平均値±SEMを表す。統計的有意性は対にしないスチューデントのt検定で決定した: *P < 0.05; **P < 0.01; ***P < 0.005; ***P < 0.001; ns = 有意でない。
全身的なベタネコール投与(BETH:400μg/mLの飲料水)は、水分消費量に影響を与えることなく、HCC(RIL175)の腫瘍負荷を増加させた(図s3A、B)(図s3C)。ベタネコールへの直接曝露はRIL175の増殖を促進しなかったことから(図s3D)、間接的なコリン作動性メカニズムが示唆される。ACh活性は全身の炎症反応に影響を及ぼすので(22, 37)、C57BL/6背景の年齢をマッチさせたRag1KOマウスとともに、独立したC57BL/6コホートで肝細胞がんモデルを繰り返した。C57BL/6マウスではベタネコールがRIL175腫瘍の増殖を促進したが、成熟した適応免疫を持たないRag1KOマウスではベタネコールは腫瘍負荷を変化させなかった(図2C)。Rag1KO-H2OとRag1KO-BETHの肝臓は同程度の骨髄蓄積を示したことから(図s3E)、ACh適応免疫メカニズムが示唆された。
図S3
図をダウンロード
新しいタブで開く
図S3
ベタネコール処理による肝腫瘍増殖の変化
(A)肝内注射から21日後のH2OおよびBETHマウスにおけるRIL175腫瘍重量。(B)(左-右)Aのマウスコホートの体重、腫瘍のない肝臓の重量、および腫瘍:健康な肝臓の比率。(C)H2OまたはBETH(ベタネコール:飲料水中400μg/mL)ケージにおける3つの異なる時点での水消費量。D)RIL175細胞への48時間ベタネコール処理後の比色MTTアッセイの結果。データは培地ウェルコントロールに対して正規化した。(E)Rag1KOマウスのフローサイトメトリーで肝臓の骨髄系免疫プロファイルを評価した独立したBETH研究のデータ。細胞数は組織重量で正規化。統計的有意性は、対にしないStudentのt検定、またはポストホックDunnett検定(D)を用いた一元配置分散分析によって決定した。棒グラフは平均値または平均値±SEMを表す: *P < 0.05; ***P < 0.005; ****P < 0.001, ns = 有意ではない。
次に、迷走神経切断モデル内の免疫組成をプロファイリングし、推定上の適応免疫参加者を同定した。肝迷走神経切断は、肝細胞癌モデルマウスの肝臓および腫瘍標本において、抗腫瘍免疫プロファイルを有意に促進した(図2Dおよび図s4A、B)。対照的に、脾臓免疫細胞の頻度はHVマウスとSVマウスでほぼ同等であり(図s4C)、肝臓特異的なコリン作動性調節が強調された。
図S4
図をダウンロード
新しいタブで開く
図S4
肝迷走神経切断は肝免疫プロファイルを変化させる
(A)フローサイトメトリーで評価した肝免疫プロファイル、細胞数はRIL175腫瘍発生後21日目の組織重量で正規化、n=10 SV、9 HV。(B)フローサイトメトリーで評価した腫瘍および(C)脾臓免疫細胞組成の平均頻度(生細胞%)。(B16-F10(n=9SV、8HV)または(E)A20モデル(n=15SV、15HV)の腫瘍発生から21日後の(D)(左-右)CD4+T細胞(CD3+CD4+)、CD8+T細胞(CD3+CD8+)、マクロファージ(CD3-CD11b+F480+)の生細胞数。Eのマウスは2つの独立したコホートから得られた。(F)RIL175のSVおよびHV肝臓におけるTreg細胞(CD3+CD4+FoxP3+)の平均頻度。A-CおよびFのマウスは同じコホートから得られた。(G)抗CD8+ T細胞の枯渇をフローサイトメトリーで確認し、代表的なフロープロットを報告し、HV肝臓における頻度(生細胞%)を報告した。棒グラフは平均値±SEMを表す。統計的有意性は対にしないスチューデントのt検定で決定した:報告値0.05<P<0.1, *P < 0.05; **P < 0.01; ****P < 0.01; ns = 有意でない。
フローサイトメトリー解析の結果、HV肝ではB細胞、樹状細胞(DC)、マクロファージ、および単球型(mMDSC)と顆粒球型/多形核型(pMDSC)を含む骨髄由来抑制細胞(MDSC)が減少していた。対照的に、HV肝ではCD3+ T細胞の増加がみられ(図2D)、これは転移B16-F10およびA20肝でも維持されているHVの特徴である(図s4D, E)。腸-肝臓の相互作用は、Treg(T制御細胞)の蓄積を制御することが報告されているが(25, 29)、脳-肝臓の破壊は、フローサイトメトリーによる肝Treg(CD4+Foxp3+)の存在量を変化させなかった(図s4F)。CD8+T細胞のみが、頻度および絶対数として評価したところ、HV肝臓内に有意な蓄積を示した(図2D、E)。
次に、CD8+ T細胞の抗体枯渇を利用して、CD8+ T細胞の関与を確認した(図s4G、図2F)。迷走神経切断術は、IgGアイソタイプコントロールの範囲内でHCC腫瘍負荷を減少させたが、CD8+ T細胞枯渇はHV腫瘍コントロールを消失させた(図2G)。これらの結果から、肝癌の迷走神経依存性腫瘍制御には、ムスカリンAChシグナルとCD8+ T細胞の両方が必要であることが示された。HV腫瘍制御が特定のCD8+ T細胞サブセットに影響を及ぼすかどうかを明らかにし、正確なコリン作動性-CD8+ T細胞相互作用を同定するために、転写研究およびin vitro研究を利用した。
コリン作動性破壊はCD8+ T細胞のエフェクター活性を促進する
同所性のRIL175 HCC腫瘍を持つマウスから得た腫瘍サンプルとマッチした(プールされた)SV肝臓3個とHV肝臓3個からソートされた(Live+CD45+)免疫細胞についてscRNA-Seq解析を行った(補足ファイル3)。厳格な品質管理の後、32,722個の細胞が下流の解析に利用された(サンプル間で平均したUMI中央値5,980)。教師なしクラスタリングでは、骨髄系、B細胞、NK、ILC1など13のサブセットが出現し(図3Aおよび図s5A-D)、scRNA-seqの結果は先のフローサイトメトリー解析をほぼ反映していた(図2D)。HVマウスは肝NKおよびILC1集団(c12_NKおよびc13_ILC1、図s4C)の中程度の拡大を示したが、これは(38)で報告された別個の遺伝子シグネチャーによって特徴づけられた(図3A、図s5A-Dおよび補足ファイル3)。
図S5
図をダウンロード
新しいタブで開く
図S5
scRNA-Seqで明らかになったHVリンパ球のプロフィール
(A)全肝臓(n = 3 / グループ)およびマッチさせたプールRIL175腫瘍サンプルのUMAPと免疫クラスター、SVおよびHV免疫(ソートされたLive+CD45+)細胞のデータは補足ファイル3も参照。(B)免疫UMAP内のサブクラスターを同定するために使用した重要な免疫マーカーの発現。(C)肝臓(腫瘍隣接)および腫瘍(腫瘍浸潤)サンプルの骨髄系、B、NK、ILC1細胞クラスター内の全細胞の割合。(D)組織(L=肝臓、T=腫瘍)および迷走神経切断の状態(H=肝迷走神経切断、S=偽迷走神経切断)別に同定したT細胞UMAP(Live+CD3+サブセット)。(E)T細胞UMAP内のサブクラスターの同定に用いた重要な免疫マーカーの発現と(F)ヒートマップ 補足ファイル3も参照。
図3
図をダウンロード
新しいタブで開く
図3
AChシグナルはCD8+抗腫瘍反応を促進する
(A) (上) SVおよびHV RIL175コホートからの肝臓(n = 3 /群)および(プールされた)腫瘍サンプルのマッチングに対するT細胞UMAP(Live+CD45+CD3+)。(下)肝臓(腫瘍隣接)および腫瘍(腫瘍浸潤リンパ球)サンプルからのscRNA-Seq教師なしT細胞クラスター内の全細胞の頻度。(B)フローサイトメトリーによる蛍光強度中央値(MFI)で測定した肝リンパ球からの生体外細胞内サイトカインおよび細胞傷害性マーカー。肝臓サンプルはRIL175腫瘍発生21日後に採取し、染色前にPMA/イオノマイシンで4時間刺激した。(C)RIL175腫瘍モデルの肝臓(左)および脾臓(右)におけるCD8+TNFα+集団の代表的なゲーティング。(D)IL2(100ng/mL)またはCD3/CD28 Dynabeads(3:1T細胞:Dynabeads)刺激下で48時間培養した後、脾臓細胞をACh曝露(2時間)+PMA/イオノマイシン活性化+ACh曝露(4時間)した。CD8+T細胞におけるTNFα+およびIL10+染色のフローサイトメトリー(n = 3 / テクニカルレプリケート)、MFIを報告。(E)scRNA-Seqリンパ球クラスター(Live+CD45+CD3+)内のAChR発現陽性率。 F)CD8+T細胞肝サブクラスターのモノクルトラジェクトリー解析(n = SV 3、HV 3)。(G) SVおよびHVサンプルからのAChR+ CD8+ T細胞のナイーブサブクラスターとエフェクターサブクラスターの比率。(H)SVおよびHVサンプルのCD8+ T細胞サブクラスターの頻度。E-Hのデータは、Aで報告した生物学的サンプルからのものである。 I)コリン作動性抗細胞傷害性CD8+ T細胞モデル。ACh遮断(例えば迷走神経切断)は、AChR依存的な細胞傷害性CD8+ T細胞サブセットの変化(HVサンプルで増加)を防ぎ、エフェクター集団を維持する。(J)脾臓OT-1 CD8+T細胞をB16-F10-OVA腫瘍細胞と12時間共培養し、エフェクター対腫瘍標的比を増加させた後の細胞傷害性アッセイ。ACh(1μg/ml)または培地対照のフローサイトメトリー分析で、最大殺傷率は熱殺傷対照を用いて決定した(n = 3 / テクニカルレプリケート)。(K) CD8creM3Rf/fマウスおよび同腹仔コントロールにおけるRIL175肝内注射8日後の代表的なIVIS® Spectrum in vivoイメージング。(L) 4時間のPMA/イオノマイシン刺激後のCD4+およびCD8+ T細胞内のCD8+GB+染色。(M)14日目に測定したRIL175腫瘍重量、四角=雄マウス、丸=雌マウス(n = 11 / 群)。データは2つの独立したコホートからプールした。特に断りのない限り、統計学的有意性は対にしないStudentのt検定、一元配置分散分析、ポストホックダンネット検定(C)または二元配置分散分析(J)により決定した: *P < 0.05; **P < 0.01; ***P < 0.05; ****P < 0.001; ns = 有意ではない。棒グラフとDは平均値を表し、報告されたひげは±SEMを表す;GB = granzyme B。
肝臓と腫瘍のサンプルをまとめてバッチ補正した後、UMAP(Uniform Manifold Approximation and Projection)により、組織および迷走神経に特異的なプロフィールを持つ9つのT細胞サブクラスターが明らかになった(図3Aおよび図s5D-F)。肝臓および腫瘍浸潤リンパ球のクラスター同一性は、遺伝子シグネチャーによって定義され、先行するscRNA-Seqランドスケープ報告(図s5Fおよび補足ファイル3(36, 39))に導かれた。最大のCD4+ T細胞サブクラスターであるc3_CD4_Proliferatingは、T細胞の維持と増殖に関与するマーカーであるIl2rbとId2の高発現によって特徴付けられた(40, 41)。HV肝臓は、HV腫瘍ではなく、c3_CD4_増殖頻度の増加を示した。対照的に、c4_Tregサブセット(Foxp3高値)は腫瘍サンプル内でより多く見られた。HV腫瘍では、免疫抑制性のc4_Tregサブセットの頻度が高かったが、これはおそらく迷走神経切断後のCD4+ T細胞の全体的な増加を反映している。これらの違いは、フローサイトメトリー解析と一致する肝臓サンプル内では、あまり顕著ではなかった(図s4F)。残りのCD4+ T細胞サブクラスター、c5_CD4_Naïve-Junとc11_CD4_Naïve-Ccl5は、高いCcr7、Lef1、Sell発現を特徴とするほぼ同等の遺伝子シグネチャーを示し、以前に報告されたクラスターアノテーション(39)と一致する。NKT細胞(c9_NKT)はHV腫瘍内で有意に増加した。肝迷走神経切断術が肝免疫の状況を著しく変化させる一方で、さらなる転写解析はCD8+ T細胞のサブセットに焦点を当てた。
ナイーブな転写シグネチャーを持つCD8+ T細胞(c7_CD8_ナイーブサブクラスター)がHVおよびSV肝サンプルで観察された(図3A)。優勢なCD8+ T細胞サブクラスターであるc6_CD8_Cytotoxicは、高いグランザイム発現、特にGzmaとGzmbの遺伝子シグネチャーを示した(図s5FとSupplemental File 3)。実際、c6_CD8_Cytotoxicクラスターは、HV腫瘍のCD45+免疫ランドスケープにおいて最大の割合(>25%)を占めていた。消耗マーカーTox(胸腺細胞選択関連高移動度グループボックス)の高発現を特徴とするc6_CD8_Exhaustedサブセットも、HVマウスでは高発現であった。このサブセットは中程度の細胞毒性遺伝子発現(Gzma、Gzmb)を示した。疲弊したCD8+ T細胞も後期分化したエフェクターメモリーCD8+ T細胞もToxを発現しているので(42)、このサブセットにはエフェクターと疲弊した特性が混在している可能性が高い。最小のCD8+ T細胞クラスターであるc10_CD8_Activatedは、HVとSVのサンプル間でほぼ同等であり、初期T細胞活性化のマーカーであるCd69の発現が最も高いという特徴があった(36)。まとめてみると、肝迷走神経切断術は、CD8+ T細胞のエフェクター特性の上昇を特徴とする抗腫瘍適応的特徴を促進した(図3A)。フローサイトメトリーや転写解析によって、HV T細胞コンパートメントの機能的プロファイリングが得られたが、その後の研究で機能的活性が検証された。
エフェクター機能を調べるため、肝細胞腫マウスの肝浸潤リンパ球を、報告されているように生体外刺激(PMA/イオノマイシン)した(36)。HVリンパ球全体で、炎症性サイトカインIFNγとTNFαの発現頻度と平均蛍光強度(MFI)の増加が観察された(図3B、図s6A)。対照的に、抗炎症性サイトカインIL10は肝迷走神経切断によって変化しなかった。転写細胞傷害性プロファイルを反映して、CD8+ T細胞はパーフォリンの発現も増加した(図3B)。一致した脾臓リンパ球の生体外分析では、SVおよびHV RIL175モデルでCD8+ T細胞の活性化が同程度であることが明らかになり(図3Cおよび図s6B)、臓器特異的な迷走神経調節が確認された。
図S6
図をダウンロード
新しいタブで開く
図S6
T細胞刺激により、HVリンパ球の応答が明らかになった
(A)フローサイトメトリーで測定した生体外肝リンパ球におけるサイトカインおよび細胞傷害性マーカーの頻度。RIL175腫瘍発生21日後に採取し、PMA/イオノマイシン下で4時間刺激したサンプル(n = SV 10、HV 9)。(B)一致した脾臓CD8+T細胞サンプルからのサイトカインおよび細胞傷害性マーカーの頻度(上)とMFI(下)。(C)B16-F10モデルにおけるCD8+ T細胞の細胞内サイトカイン染色(IFNγ、TNFα)に続いて報告されたMFI。AおよびBで報告した刺激および染色。 (D) 肝A20-腫瘍組織からのIL2サイトカイン分析、データは肝組織重量で正規化(n = 7 SV, 8 HV)。Dにおけるサイトカインレベルは、カスタマイズされたProcartaplex Luminexイムノアッセイによって決定された。棒グラフは平均値±SEM。統計的有意性は対にしないスチューデントのt検定によって決定した: 0.05<P<0.1;*P<0.05;**P<0.01;***P<0.005;ns=有意ではない。
転移性肝モデルでもHV誘発炎症マーカーは同等であり(図s6C, D)、強固な肝神経免疫アークが強調された。コリン作動性の破壊が抗腫瘍免疫応答を促進する一方で、我々は次にACh活性がCD8+ T細胞に直接影響を及ぼすかどうかを確認しようとした。
AChシグナルはCD8+ T細胞の抗腫瘍活性に影響する
AChとCD8+ T細胞の直接的な相互作用を評価するために、ACh急性曝露(4時間)後の細胞内サイトカイン産生を、記載されているように測定した(27, 36)。脾臓リンパ球は、報告されているようにIL2またはCD3/CD28で拡大した(43)。ACh曝露は、CD8+T細胞のTNFα産生を用量依存的に減少させたが、IL10は減少させなかった(図3D)。これは、AChがマクロファージのサイトカイン放出を抑制するという先行報告(27、28)を反映している。我々は、AChで処理したCD4+ T細胞内で同様のサイトカイン調節を観察し、広範なリンパ球制御を示した(図S7A)。
図S7
図をダウンロード
新しいタブで開く
図S7
コリン作動性シグナルはT細胞の炎症を抑制する
(A)IL2(100ng/mL)またはCD3/CD28 Dynabeads(3:1T細胞:Dynabeads)刺激下で48時間培養した後の脾臓CD4+ T細胞集団。増加するACh濃度に曝露したCD4+ T細胞におけるPMA/イオノマイシン活性化後のTNFα+およびIL10+細胞内サイトカイン染色、MFI(上)および親の頻度(下)を報告、n=3テクニカルレプリケート。さらなる刺激手順はExpanded Methodologyで報告。(B)ベタネコールまたは(C)4-DAMP(1,1-ジメチル-4-ジフェニルアセトキシピペリジニウムヨージド:AChR M3アンタゴニスト)に暴露したリンパ球のTNFα+細胞内染色。Aのプロトコールに従った刺激手順、n = 3テクニカルレプリケート。(D)RIL175およびB16-F10腫瘍細胞株の48時間ACh処理後のMTTアッセイ結果。MTTアッセイは細胞の生存率と増殖の代用となる。データは培地ウェルコントロールに対して正規化した。(E)ムスカリン性およびニコチン性AChR遺伝子の発現によって決定されたAChR+およびAChR-サブセットのバイオリンプロット。AChR+細胞(点線上)は、複合AChR発現>0.05として同定され、クラスターは組織(L=肝臓、T=腫瘍)および迷走神経切断の状態(H=肝迷走神経切断、S=偽迷走神経切断)によって分けられ、n=3/群(肝臓)またはマッチさせたプール(腫瘍)。統計的有意性は、対にしないStudentのt検定(B)またはポストホックDunnettの検定(A、C、D)を伴う一元配置分散分析(one-way ANOVA)によって決定した。非バイオリンプロットのデータは平均値を示し、表示されたひげは平均値±SEMを示す: *P < 0.05; **P < 0.01; ***P < 0.005; ****P < 0.001, ns = 有意ではない。
次に、ムスカリンAChRの関与がCD8+ T細胞の調節に直接関与しているかどうかを調べた。迷走神経切断によるACh枯渇はCD8+TNFα+発現を促進したが、ベタネコール処理はex vivo脾臓T細胞の細胞内TNFα産生を減少させた(図s7B)。対照的に、CHRM3を優先的に標的とする選択的AChRアンタゴニストである4-DAMP(1,1-ジメチル-4-ジフェニルアセトキシピペリジニウムヨージド)への暴露は、刺激されたCD8+T細胞内で用量依存的にTNFα+発現を促進した(図s7C)。これらの所見はACh-腫瘍活性を直接否定するものではないが、ACh暴露(48時間)はin vitroでの腫瘍細胞株の増殖や生存率に影響を与えなかった(図s7D)。
肝迷走神経切断は肝AChレベルを減少させる(図2A)。興味深いことに、scRNA-seq解析により、ほとんどのHVリンパ球でAChR+サブセットが増加していることが明らかになり、迷走神経切断後の代償的なアップレギュレーションが示唆された(図3Eおよび図s7E)。その後のパスウェイ濃縮解析により、サブセット特異的なコリン作動性影響が同定された。ACh-免疫活性に注目するため、肝組織で発現したコリン作動性活性と炎症性活性化を報告する遺伝子オントロジー(GO)パスウェイを含む「神経免疫パネル」(補足ファイル4)を作成した。予想通り、HV肝臓および腫瘍(Live+CD45+)細胞は、免疫応答に関与するリンパ球活性化(GO:0002285)および神経炎症応答(GO:0150076)を含む、濃縮された炎症性応答を発現した(図S8A, Bおよび補足ファイル4)。
図S8:
図をダウンロード
新しいタブで開く
図S8:
scRNA-Seqにより、CD8+ T細胞クラスターにおける異なるACh制御が同定された。
(A)神経免疫パネルとして12のパスウェイを選択した(補足ファイル4参照)。肝迷走神経切断肝臓と偽迷走神経切断肝臓、それぞれLHとLSにおける神経免疫パネルのパスウェイ濃縮解析(左)。(B)(右)肝迷走神経切断腫瘍および偽迷走神経切断腫瘍、それぞれTHおよびTSにおける神経免疫パスウェイの濃縮解析。統計的に有意なパスウェイのみがドットプロットに表示され、色はPadjを示し、サイズはGeneRatioを反映する。すべての免疫(ソートされたLive+CD45+)細胞のデータ。(C)(左-右)図3Aを参照したCD8+T細胞クラスターにわたる神経免疫パスウェイの濃縮解析:c7_CD8_Naïve、c10_CD8_Activated、c6_CD8_Cytotoxic、c8_CD8_Exhausted。scRNA-Seqデータについては、n = 3 / 肝臓とマッチしたプール腫瘍サンプルのグループ。
すべてのHV CD8+T細胞サブセットは、濃縮された神経炎症性発現を示したが(図s8B)、ACh応答(Cellular Response to Acetylcholine [GO:1905145]は変化した、図s8C)。残りのCD8+ T細胞サブセットとは対照的に、c10_CD8_Activatedサブセットのみが、迷走神経切断マウスにおいてACh細胞応答を豊富に示した。対照的に、より豊富なc6_CD8_Cytotoxicサブセットとc8_CD8_Exhaustedサブセットは、SVサンプル内でACh細胞応答の上昇を示し、AChはサブセット特異的相互作用を示すことが示唆された。
この提案は、肝臓と腫瘍のCD8+ T細胞の時系列解析によって裏付けられた。Monocle3(44)を用いた擬似時間解析と軌跡解析により、CD8+ T細胞が軌跡経路を横切ってダイナミックに分岐していることが明らかになった(図3F)。c6_CD8_Cytotoxicサブセットだけが、排出されたクラスターへの軌道分岐を含んでいた。実際、肝臓と腫瘍サンプルのAChR+ T細胞クラスターの比率は、SVマウスとHVマウスで、活性化サブセット:ナイーブサブセットの比率が同程度であったのに対し、HVサンプルでは、細胞傷害性:活性化サブセットが増加し、疲弊性:細胞傷害性サブセットが減少した(図3G)。AChRアゴニストモデル(図2B)、in vitro研究(図3D)、およびトランスクリプトーム解析(図3E-Hおよび図s8)を総合すると、AChの破壊(例えば、迷走神経切断による)はAChRシグナルを遮断し、AChRシグナルを介してCD8+ T細胞の細胞傷害性機能の維持と拡大を促進することが示唆される(図3I)。
腫瘍環境における細胞傷害性のACh調節を検証するために、MHC-Iクラス制限のあるオバルブミン由来ペプチドに特異的なCD8+T細胞を発現するトランスジェニックOT-1マウスからCD8+T細胞を単離し、OT-1 CD8+T細胞をB16-F10-OVA腫瘍細胞と共培養した。ACh投与はB16-F10-OVA細胞のCD8+ T細胞溶解を阻害した(図3J)。
AChはムスカリン性AChR依存性のシグナル伝達を介してCD8+ T細胞のエフェクター機能を強力に抑制する。アンタゴニスト研究(図7C)と転写解析(図9A)は、末梢腫瘍細胞増殖の制御因子(21, 22, 45)であるムスカリン性CHRM3受容体が、CD8+ T細胞の細胞傷害機能の調節に寄与していることを示唆している。そこで、Cre/lox操作によりCD8+T細胞上のChrm3発現を欠損したマウス(CD8creM3Rf/f)を作製した。同腹のコントロールマウスと比較して、CD8creM3Rf/fマウスは、非吻合マウスにおいて肝細胞癌の増殖がわずかに減少した(図3Kおよび図s9B)。注目すべきは、CD4+ T細胞ではなく、CD8creM3Rf/f肝CD8+ T細胞が、細胞傷害活性のマーカーであるグランザイムB発現の増加を示したことである(図3L)。CD8creM3Rf/fマウスは、同腹のマウスと比較して腫瘍負担が有意に小さかったが(図3M)、遺伝子修飾は体重、非腫瘍肝臓重量を損なわず、総リンパ球蓄積量も変化させなかった(図S9C、D)。
図S9:
図をダウンロードする
新しいタブで開く
図S9:
AChRシグナル伝達は肝腫瘍増殖を変化させ、CD8+ T細胞の細胞傷害性に影響を及ぼす
(A)示差的発現遺伝子(DEG)scRNA-Seq解析;RIL175モデルにおけるAChR+肝T細胞とAChR-肝T細胞のボルケーノプロット(n = 3 /群)。縦点線は平均log2倍変化(log2FC)に基づく1.5倍変化を示し、横点線は0.05 Padjカットオフを示す。(B)ルシフェラーゼ発現RIL175腫瘍の肝内注射8日後の代表的なin vivoイメージング(IVIS® Spectrum)(n = 8 CD8creM3Rf/fおよび7同腹子コントロール)。(C)(左-右)肝臓内注射から14日後のCD8creM3Rf/fおよび同腹子コントロールにおける総体重、非腫瘍性肝臓重量、およびRIL175腫瘍:体比(n = 11 /群)。D)CD8creM3Rf/fおよび同腹仔コントロールにおけるCD4+およびCD8+ T細胞の頻度(生細胞率)。B-Dにおける統計的有意性は、対応のないスチューデントのt検定によって決定した。棒グラフは平均値±SEMを表す:0.05<P<0.1; *P < 0.05; 1, ns = 有意ではない、または報告値。
まとめると、この研究から、ダイナミックな迷走神経➔肝軸が明らかになった。肝ACh活性は、肝CD8+T細胞サブセットに多様で強固な調節をもたらし、特にCHRM3シグナル伝達を介して細胞傷害性応答に影響を及ぼす(図3)。コリン作動性の変化だけでなく、肝臓がんはマウス(図s8)や男性(図s2A)において神経関連経路を破壊する。臓器間のクロストークは本質的に双方向的であり(15, 16)、迷走神経➕脳弓が肝がんに関連した神経学的副作用を形成している可能性が高い。次に、肝迷走神経切開が肝がんに関連する行動(異常な疲労など)を変化させるかどうかを調べた(46-49)。
迷走神経切断はHCC腫瘍マウスにおいて神経学的機能を改善する
施設馴化後、迷走神経切断HCCマウスは、表現型、オープンフィールドテスト(OFT)、Y迷路(図4A)など、確立された一連の行動テストを受けた(48、50、51)。迷走神経切断と腫瘍によって誘発される神経学的行動に対処するため、RIL175腫瘍移植の前に一連の「ベースライン」試験を行った。後者のタイムポイントでは、事前のin vivoイメージングに基づき、SVマウスとHVマウスの腫瘍負荷がより小さく同程度であった時期(早期)と、腫瘍がより大きく腫瘍の変動が大きかった時期(後期)の行動を評価した(図1C)。運動は、以前に報告された交感神経-腸管免疫機構(52)を介して肝腫瘍負担を減少させたが、適度な運動負荷試験はHV腫瘍制御に影響を与えなかった(図S10A)。
図S10
図をダウンロード
新しいタブで開く
図S10
迷走神経シグナル伝達の障害が行動を形成する
(A)肝内注射から26日後の行動モデルにおけるRIL175腫瘍の重さ。(B)ベースラインおよび後期タイムポイント(n = 3 /群、10分間隔で記録)における日中追跡中のフェノタイパー距離(中心点からの総移動距離)、図4A参照。(C)ベースライン(膣切開後)、早期(腫瘍摘出後)、後期(腫瘍摘出後)のタイムポイントにおけるY字迷路での活動性評価(総移動距離、平均速度、総不動時間を含む)。(D)OFT中のベースライン(膣切開後)および後期(腫瘍摘出後)のタイムポイントにおける活動性評価(総移動距離、平均速度、総無動時間を含む)、およびOFZ内と周辺部での時間の比率(中心:端)。(E)終点腫瘍重量と後期活動の線形相関: Y字迷路アームへの進入時間(左)とOFZ時間(右)。パネル内のすべてのマウスは同じ行動研究から得られたもので、n = 8 /群。統計学的有意性は、線形回帰(E)または対応のないスチューデントのt検定により決定した、OFT=オープンフィールドテスト、OFZ=オープンフィールドゾーン。棒グラフは平均値±SEMを表す:報告値0.05<P<0.1, *P < 0.05; **P < 0.01; ns =有意ではない、または報告値。
図4
図をダウンロード
新しいタブで開く
図4
迷走神経シグナル伝達の障害が神経認知反応を形成する
(A)施設馴化、ベースライン(膣切開後、腫瘍なし)、早期(腫瘍後)、後期(腫瘍後)のタイムポイント、72時間のフェノタイパー追跡、Y迷路、オープンフィールドテスト(OFT)テストを含む行動解析の概略図。(B)(左)ベースライン(上)と後期(下)の12時間の平均のフェノタイパーの距離(中心点からの移動距離)。(右)ベースライン(上)と後期(下)の12時間のタイムポイントにおける不動性(睡眠/活動性)の追跡。(C)ベースライン(上)と後期(下)のタイムポイントにおけるフェノタイパーの夜間距離、ポイントは10分間隔を示す。PhenoTyper 3000ケージが利用可能であったため、フェノタイピング実験が制限され、残りのマウスはフェノタイピング実験中ずっとシングルハウジングに入れられた。(D)SVおよびHVマウスにおける72時間追跡の平均値、ベースラインおよび後期タイムポイントのペア。(E)ベースライン、初期、後期の各タイムポイントにおけるY字迷路の交替率(左)と全アーム進入数(右)(5分間の自由探索)。(F)(左)ベースラインと後期のタイムポイント(30分間の自由探索)におけるオープンフィールドゾーン(OFZ)での総滞在時間。(右)SVとHVの後期タイムポイントにおける代表的なOFT追跡プロット。B-Fのマウスは同じコホートからのもの。(G)非腫瘍(NT)および腫瘍(T)のセカル微生物叢移植(CMT)レシピエントと行動解析のタイムライン: TドナーはPBS/マトリゲルマトリックス中のRIL175細胞の肝内注入を受け、NTドナーはPBS/マトリゲルマトリックスビヒクルの肝内注入を受けた。21日目に回収され、プールされた濾過された内容物は、ポリエチレングリコール(PEG)下剤処理後、4回の経口経口投与(100μL)によりNTまたはTレシピエントに移植された。ベースライン(cecal microbiota移植前=CMT前)およびCMTの時点で実施したフェノタイパーおよびOFT試験。(H)CMT試験中のフェノタイパーの可動性(中心点からの移動距離の合計)と12時間の平均(n = 7 NT、5 T)。(I)12時間平均でマウスから測定した日中の不動性。日中の時間を示すすべての表現型データのハイライトオーバーレイ。(J)肝内注射を受けたRIL175腫瘍マウス(RIL175)または非腫瘍対照マウス(CON)の脳幹部の代表画像。統計的有意性は、非対または対Student's t testで決定した:(報告値)0.05<P<0.1, *P < 0.05; **P < 0.01; ns =有意でない、または報告値を記載。ひげは±SEM、棒グラフは平均を示す。
まず、PhenoTyper 3000ケージに収容したマウスの疲労を評価し、日中(休息時)および夜間(移動時)の活動と無動状態を把握した(50)。膣切開したマウスは、特に夜間のサイクルにおいて、移動性(移動距離)と不動性(睡眠/活動性)によって測定される疲労の減少を示した(図4B-Dおよび図s10B)。肝臓がんはSVマウスとHVマウスの疲労を増悪させたが、HVマウスは睡眠・活動時間を短く維持した(図4B-D)。
運動以外にも、認知機能と不安様行動をそれぞれY字迷路とOFT(48, 51)で評価した。各Y字迷路評価において、SVマウスとHVマウスは正常な自発交代行動を示し(図4E)、探索と急性空間記憶に関する認知能力は同等であることが示唆された(48)。後期時点では、HVマウスはY字迷路の腕の総進入距離、総移動距離、平均速度、不動性で評価した移動性の増加を示した(図4Eおよび図s10C)。移動性の増加は、ベースラインではなく、後期のOFT試験でも観察された(図s10D)。肝腫瘍の発生に伴い、HVマウスは不安様行動の減少を示し、オープンフィールドゾーン(OFZ)探索の増加によって示された(図4Fおよび図s10D)。要約すると、腫瘍を有するHVマウスは神経学的転帰が改善し、特に疲労の軽減、運動能力の向上、不安関連行動の改善が認められた。
行動はSVとHVの腫瘍負担の違いを反映している可能性があるが、最終腫瘍重量はY字迷路やOFTの活性とは相関しなかった(図s10E)。肝迷走神経完全性は肝腫瘍負荷を形成したが(図1D-F)、SV腫瘍サイズは行動障害の有意な促進因子ではなかった(図s10E)。その結果、全身性の肝細胞増殖異常症が神経学的転帰を形成するという仮説が立てられた。興味深いことに、非肝迷走神経による行動の調節は、腸-迷走神経-脳の相互作用の文脈で報告されている(17)。迷走神経は主に求心性線維で構成され、消化管(GI)を網羅し、微生物の合図に反応する(17, 53)。その結果、HCCマウスの腸内細菌叢ががん関連行動を形成するのに十分であるかどうかを調べた。
プールされたRIL175担癌ドナー(T)または腫瘍のない肝内外科的コントロール(NT)から経口経口投与で、腸内細菌叢移植(CMT)を行った(図4G参照)。CMTの前と後に、表現型解析とOFT試験を行った(図4H)。TレシピエントとNTレシピエントは同程度のOFT探索と活動性を示したが(図s11A、B)、表現型解析により、CMT後のTレシピエントでは、移動性(移動距離)の低下と不動性(睡眠/不活動性)の増加が明らかになった(図4H、Iおよび図s11C-E)ことから、HCC肝臓からの腸内細菌叢異常は、がん関連疲労を促進するのに十分であることが示唆された。迷走神経切断マウスにおける以前の行動解析(図4B-D)とは対照的に、CMT研究では、主に日中(休息期:図4I)に疲労行動が変化することが報告され、おそらく反復CMTによる概日リズムの乱れと関連していることが示唆された。
図S11
図をダウンロード
新しいタブで開く
図S11
腫瘍を有するCMTは運動量を変化させる
(A)T(腫瘍)およびNT(非腫瘍)の食道微生物叢移植(CMT)研究のOFT結果(図4G参照)。CMT前(上)とCMT後(下)のOFZ総時間とOFZと周辺領域で過ごした時間の比率(中央:端)。(B)CMT前(上)とCMT後(下)のOFT活動(総移動距離、平均速度、総無動時間を含む)、n=10NT、8Tマウス。(C)CMT前の一部のマウス(n = 4 /群)におけるフェノタイパー距離(中心点からの総移動距離)と(D)無動時間(睡眠/活動時間)。2日目にソフトウェアの不具合があったため、1回目と3回目のデータのみを報告。PhenoTyper 3000のケージはCMT実験のnサイズを決定し、残りのマウスは表現型解析の間単独でケージに入れられた(E)CMT後の試験(n = NT 7, T 5)の日中および夜間のフェノタイパー距離(中心点からの移動距離の合計)。(F)腫瘍発生から14日後に染色したCONマウスとRIL175マウスのNTS領域あたりのcFOS数(n = 4 NT, 5 T)。F以外は同じCMT研究のS9パネルのマウス;OFT=open field test、OFZ=open field zone。グラフは平均値±SEM。統計的有意性は対にしないスチューデントのt検定で決定した:ns = 有意でない。
これらの結果から、腸内微生物はがん関連行動を変化させるのに十分であることが示された。前述のように、消化管は迷走神経求心性神経に強く支配されている。迷走神経求心性神経は、脳幹内の孤束核(NTS)に情報を伝達する感覚線維である(29)。非腫瘍の対照マウスと比較すると、HCCマウスはc-Fosの発現によって定義されるように、NTSの適度な活性化を示した(図4Jおよび図s11F)。これらの所見は、がんに関連した腸内環境の異常が脳の特徴に影響を与えていることを示している。しかし、迷走神経切断マウス(Baseline)とCMTレシピエントはともに、疲労パターンに変化を示した(図4B-D)。その結果、腸内細菌叢の異常が迷走神経切断や肝臓がんに起因するものであるかどうかを調べた。
迷走神経切断ではなく腫瘍が糞便微生物叢組成を強固に形成する
迷走神経活動は消化管通過時間に影響を及ぼし、それ自体が腸内細菌叢組成の主要なドライバーである(54)。肝枝(横隔膜下)迷走神経切断術は胃排出とGI通過を障害するため、幽門形成術を併用する必要がある(24、45、55)。しかし、正確な肝枝迷走神経切断術では、胃通過時間を変化させることはできなかった(図s12A)。実際、迷走神経切断を行った非腫瘍群(HVx)と偽コントロール群(SVx)は、16S rRNA-Seqで評価した糞便中の腸内細菌叢組成とα多様性は同等であった(図s12B、C)。実際、迷走神経切断の状態よりもケージ効果の方が、腸内細菌叢プロファイルに大きな影響を与えた(図s12Bおよび補足ファイル5)。
図S12
図をダウンロード
新しいタブで開く
図S12
抗腫瘍免疫と迷走神経切断されたマイクロバイオームとの関連は認められない
(A)カルミンレッド試験で測定したGI(消化管)通過時間。(B)肝迷走神経切断術(HVx)または偽手術(SVx)を受けた非腫瘍マウスの糞便微生物叢サンプルのβ多様性(Jaccard、Weighted UniFrac)およびα多様性(FaithのPD、Shannonエントロピー)。迷走神経切断後、マウスは無作為に3週間同居させた。糞便サンプルの16S rRNA-Seq後に決定されたJaccard(左)およびWeighted UniFrac(右)プロット。(C)SVxマウスとHVxマウスにおける細菌組成(次数)。A-Cのマウスは同じコホートのもので、補足ファイル5も参照。(D)フローサイトメトリーによるSVxマウスとHVxマウスの免疫プロファイリング。(E)SVxおよびHVxマウスの肝サンプル(n = 14 /群)の組織重量で正規化したLive+免疫細胞の細胞数。(F) 4時間のPMA/イオノマイシン刺激後の生体外リンパ球におけるグランザイムB、IFNγ+およびTNFα+染色、CD4+およびCD8+ T細胞について報告されたMFIおよび(G)親頻度、n = 7 /群。(H)コホージング腫瘍実験のためのケージング戦略。迷走神経切断後、HVxおよびSVxマウスを、RIL175腫瘍開始前に非飼育ケージ(SVまたはHV)または同居ケージ(cSVおよびsHVマウス)に無作為に割り付けた。腫瘍重量と最終糞便微生物叢サンプルは21日目に評価した。I-Rで報告した同居実験では、n=8SV、7cSV、8cHV、8HV。(I)担癌SV、cSV、sHV、HVマウスにおける同居終了前の消化管通過時間をcarmine red testで評価。(J)SV、cSV、cHV、HVの糞便サンプルのJaccard β多様性プロットとシャノンエントロピー(α多様性)の報告。(K)KからのJaccard距離、q値を報告したペアワイズPERMANOVA。(L)糞便サンプルからの16S rRNA-Seqにより決定された細菌組成。(M)(左)腫瘍重量および(右)非腫瘍肝重量(コホージング実験における21日時点)、色は群を示し、形状は迷走神経切断の状態を示す:丸=偽手術、四角=肝迷走神経切断。(N) (F)で報告した刺激後の肝組織における生体外細胞内サイトカイン染色。CD8+TNFα+マウスのMFI。(O)Nに示したCD8+TNFα+のMFI(左)と頻度(右)の両方を、同居群と非同居群に分けて示した。(P) 肝癌発生前(Tumor_Vagotomy = pre_SV, pre_HV)と実験終了時(Tumor_Vagotomy = post_SV, post_HV)のSVとHVの糞便サンプルの16S rRNA-Seq評価から得られたベータ(Jaccard)とアルファ(Shannon Entropy)のプロット。SVおよびHVのβ多様性組成はJ. (Q) P(pre_SVに設定)からのJaccard距離、ペアワイズPERMANOVAが実行され、すべてのq値は<0.001であった。(R)糞便サンプルからの16S rRNA-Seqにより決定された細菌組成、上位2分類群(Order)は腫瘍前後のタイムポイントで報告されている。棒グラフ(A、E-G、I、M-O、およびR)は平均±SEMまたは箱ひげ図(最小-最大ひげ)を表す: *P < 0.05; **P < 0.01; ***P < 0.005; ****P < 0.001, ns =有意ではない。
担癌マウスにおける免疫評価(図2Dおよび図3A)とは対照的に、SVxおよびHVx肝臓は、同様の免疫細胞組成(図s12D、E)およびCD4+およびCD8+T細胞における同等の炎症性サイトカイン発現(図s12F、G)を示した。これらの所見は、迷走神経切断に依存した免疫の変化には肝腫瘍が必要であることを示している。次に、肝迷走神経切開によって誘発された微生物叢異常と肝腫瘍負荷の役割をさらに確立するために、コホージング実験を利用した。
マウスのcoprophagic activityと相互グルーミングは、長期同居の間に糞便微生物叢組成を正常化する(56)。迷走神経切断術の後、無作為化したSVマウスとHVマウスのサブセットを、RIL175細胞を肝内注入する前に同居させた(それぞれcSVとcHV)(図s12H)。同居させたが、腫瘍のあるマウスの消化管通過時間は変化せず(図s12I)、通過時間は腫瘍のないマウスの結果と同等であった(図s12A)。全群で糞便微生物叢のα多様性は同程度で、同居下でのβ多様性(Jaccard)と組成(Order)はSVとHVの糞便微生物叢の中間的な表現型を示した(図s12J-Lおよび補遺ファイル5)。同居マウスは腸内細菌叢プロファイルを正常化したが、腫瘍負荷、肝臓重量、CD8+TNFα+発現は非同居マウスと同等であった(図s12M-O)。非飼育下のSVマウスとHVマウスは、迷走神経依存性の腸内細菌叢異常症を示す明確な糞便中細菌叢組成を示したが(図s12J-L)、肝臓がんは腸内細菌叢組成を著しく変化させた(図s12P)。腫瘍後のSVおよびHVの腸内細菌叢の特徴は、腫瘍発生前に採取した糞便中の細菌叢サンプルよりも類似しており(図s12P-Rおよび補足ファイル5)、迷走神経切断よりもむしろ肝細胞癌の方が、糞便中の腸内細菌叢組成に強い影響を与えることを示している。以上のことから、迷走神経により誘発された腸内細菌叢の異常は、肝CD8+T細胞抗腫瘍免疫とその後の肝腫瘍負荷を制御するのに十分ではないことが示された。
腸内細菌は迷走神経を介して肝免疫を調節する
我々の研究室では以前、腸内細菌が肝NKT蓄積に影響する腸-肝アークを介して肝腫瘍負担を調節することを報告した(33, 57)。コホージング実験(図s12)により、迷走神経切断された肝臓と腸の "直接的な "相互作用は、CD8+ T細胞の蓄積や抗腫瘍機能に影響を与えないことが示唆された。しかし、行動CMT研究(図4H、I)からは、肝臓がんの影響は「間接的な」腸内細菌叢-脳シグナルを介して媒介される可能性が示唆された。したがって、腸内細菌叢は迷走神経を介して肝免疫寛容の維持に役立ち、迷走神経障害(迷走神経切断など)は抗腫瘍免疫の拡大を促進するという仮説を立てた。
肝細胞癌に依存した微生物叢異常症を特異的に評価するために、8人のRIL-175担癌ドナー(T)と7人の非担癌コントロール(NT)からCMTを行った(図5A)。レシピエントは、ドナー特異的な糞便を含む経口摂取を繰り返す前に、偽肝迷走神経切開または肝迷走神経切開を受けた。肝免疫プロファイリングと腸内細菌叢プロファイリングの両方が行われ、CMT前と実験終了時に糞便サンプルが採取された。さらに、小腸(空腸、回腸)および大腸内容物を採取し、消化管全体の腸内細菌叢組成を調べた(図5B)。
図5
図をダウンロード
新しいタブで開く
図5
腸内微生物は迷走神経-肝臓アークを介して肝臓免疫を制御する
(A)迷走神経切断の状態(偽迷走神経切断=SVまたは肝迷走神経切断=HV)と固有のドナー(非腫瘍=NTまたはRIL175腫瘍=T)を示す。(B)レシピエントはユニークなドナーの微生物叢を受け取った。CMTの手順については「拡張された方法論」に詳述した。ドナー:n = 8 NT、7 T;レシピエント:n = 9 SVNT、10 HVNT、9 SVT、9 HVT。緑色の円は、最初の微生物叢の経口投与から10日後に16S rRNA-Seqでレシピエントの微生物叢を解析するための収集領域を示す。(C)CMTレシピエントによる小腸、セカル、および糞便サンプルの重み付けUniFrac(β多様性)解析。色=ドナー群、形=迷走神経切断の状態、大きさ=シャノンエントロピー(アルファ多様性)。(D)Cで解析した糞便サンプルのJaccard距離(グループのβ多様性)、q値を報告したペアワイズPERMANOVA。(E) 16S rRNA-Seqによって決定された糞便微生物叢組成(Order)。(F)HVNTおよびHVTの糞便微生物叢サンプルのlefSE分析(84)(クラスカル・ワリスα=0.001、ウィルコクソン検定α=0.001)、線形判別分析(LDA)スコア>2(左)およびクラドグラム(右)を介して可視化された発現差のある分類群、アスタリスクはGで報告された細菌頻度の2つを示す。(H) Partek Flowを用いて同定した蛍光ベースのクラスタリングによるフローサイトメトリーで解析したLive+CD8+肝細胞からのUMAP、(左)全CD8+T細胞のUMAP、(右)レシピエントグループに分離したUMAP。(I) フローサイトメトリーのマニュアルゲーティングにより測定した生体外細胞内サイトカイン染色。白血球刺激(PMA/イオノマイシン)および染色は図3Bおよび拡大法で報告した通り。グランザイムB+(GB+)、IFNγ+TNFα+サブセットに対するCD8+T細胞の割合。(J)(左-右)GB+、IFNγ+、およびTNFα+ CD8+ T細胞のMFI。IとJの色は固有のドナーを示し、形は迷走神経切断の状態を示す。(K)肝免疫を制御する迷走神経-肝-腸アークのグラフ表示。肝臓がんは腸内細菌叢を変化させ、その後の腸内細菌叢の異常は、(1)がん関連疲労と(2)肝迷走神経切断マウス内での抗腫瘍免疫活性を促進する。迷走神経コリン作動性シグナルは肝免疫細胞組成を調節する。迷走神経活性化の阻害は、特にCD8+ T細胞におけるムスカリンシグナル伝達を介して、抗腫瘍免疫を促進する。CD8+T細胞のエフェクター応答が拡大することで、肝腫瘍負荷が抑制される。棒グラフはTukey whisker plot(D)または平均±SEM(G、I、J)を示す。特に断りのない限り、統計学的有意性は一対のPERMANOVA(D)または一元配置分散分析とTukeyの事後多重比較(G、I、J)によって決定された: **P < 0.01; ***P < 0.005; ns = 有意でない。
NTおよびT細菌カクテルからのCMT(1:1の糞便内容物:50%グリセロール)は、レシピエントの糞便微生物叢組成を有意に変化させた(図s13A-Cおよび補足5)。小腸、盲腸、および糞便サンプルは、部位特異的で明確な腸内細菌叢組成と多様性を示した。小腸は最も低いアルファ多様性(シャノンエントロピー)を示し、腫瘍ではなく迷走神経切断の状態が細菌組成に大きな影響を及ぼした(図5C、E、図s13D、および補足ファイル5)。対照的に、糞便および便の細菌組成は腫瘍(ドナー)の状態を反映し(図5C-E、図s13D、および補足ファイル5)、糞便のJaccard距離(β多様性)は腫瘍に依存した変化を強調した(図5D)。その結果、その後の解析では、迷走神経切断マウス(すなわち、HVNT対HVT)におけるHCC腫瘍依存性変化の特徴づけに焦点を当てた。厳格なLefSe(線形判別分析[LDA]の効果量: Kruskal-Wallisα=0.001、Wilcoxonα=0.001、LDA>2)により、糞便中のHVNT微生物叢とHVT微生物叢の間に多数の細菌変化が認められたが(図5F)、HVT対SVTの解析では有意な分類群の濃縮は認められず、腫瘍依存性の変調を支持した。LefSeは小腸HVTサンプル内で濃縮された分類群を同定しなかったが、便および糞便HVT群集はともにClostridial種の増加を示した(図5F、Gおよび図s13E)。HVT群とSVT群ともに、糞便微生物叢においてClostridiales属の増加が認められたが、Bacteroidales属の増加は認められなかった(図5G)。HVTの糞便微生物叢はまた、肝臓悪性腫瘍や腸迷走神経刺激に関連する硫酸還元性分類群であるDesulfovibrionales(図5F、G)に富んでいた(58, 59)。Desulfovibrionales属の常在菌にはBilophilaとDesulfovibrioが含まれていた(図S13F)。
図S13
図をダウンロード
新しいタブで開く
図S13
迷走神経切断よりも腫瘍の状態がCMT所見を左右する
(A)腫瘍(T)および非腫瘍(NT)ドナーの凍結糞便内容物の16S rRNA-Seqによる糞便微生物叢解析、糞便微生物叢移植(CMT)研究で報告されたα多様性(Shannon Entropy)およびβ多様性(Weighted UniFrac)。(B)NTおよびTドナーのレポジトリから得られた細菌組成(Order)。(C)α(シャノンエントロピー)とβ多様性の時系列プロット:(上)Jaccard、(下)Weighted UniFracでCMT前の微生物叢とCMT後(最終)の糞便を評価。左から右へ、色は採取時点(none = ドナー、pre = CMT前の糞便サンプル、post = 最終糞便採取)、ドナーの状態(ユニークドナーCMT)、グループ(すなわち、SVNT、HVNT、SVT、HVT)を示す。CMT解析では、ドナー:n = 8 NT、7 T;レシピエント:n = 9 SVNT、10 HVNT、9 SVT、9 HVT。(D) 小腸、セカル、糞便サンプルからのJaccard β多様性とシャノンエントロピー(α多様性)、図5Cに示したWeighted UniFrac。(E)HVNTおよびHVT糞便微生物叢サンプルのLefSE解析(84)。小腸、盲腸、糞便サンプルの(左-右)クラドグラムにより発現の異なる分類群を可視化。GI管にわたるLefSE解析:α = 0.01 Kruskal-Wallis、α = 0.01 Wilcoxon検定、LDA > 2(図5Fより厳密でない)。SVTおよびHVTの糞便サンプルでは、これらのLefSe設定を用いても、濃縮された細菌分類群は報告されなかった。(F)糞便サンプルからの細菌種(左)Bilophilaおよび(右)Desulfovibrioの相対頻度。Fの統計的有意性は、一元配置分散分析(one-way ANOVA)とポストホックTukey検定によって決定した。Fの棒グラフは平均値±SEMを表す: *P < 0.05; **P < 0.01; ns = 有意でない。
微生物叢移植は、全身的な免疫調節とCD8+T細胞活性の変化に関連している(60)。続いて、肝細胞癌(TCC)の微生物叢がCD8+ T細胞の蓄積や機能を形成するかどうか、またその程度を評価するために、肝免疫の特徴をプロファイリングした。SVT肝臓とHVT肝臓の肝活性化が同等であることから、肝細胞癌の腸内細菌叢異常による直接的な腸-肝相互作用が示唆される。これとは対照的に、迷走神経-肝モデルでは、SVTマウス(迷走神経無傷)ではコリン作動性免疫抑制が維持されるのに対し、HVTマウス(迷走神経障害)ではHV肝細胞腫モデルで観察されるようにCD8+ T細胞のサイトカインおよび細胞傷害性機能の上昇が観察されるため、SVTとHVTで異なる反応が観察されるであろう(図S14Aおよび図3)。
図S14:
図をダウンロード
新しいタブで開く
図S14:
迷走神経-肝軸は肝免疫組成と機能を変化させる
(A)CMT研究における推定腸➔肝(上)および迷走神経➔肝(下)の肝反応モデル。X軸はレシピエント群、Y軸は肝炎またはCD8+T細胞のエフェクター機能を示す。T微生物叢が腸-肝相互作用を介して肝反応を引き起こすのに対して(上)、SVTマウスは迷走神経完全性、すなわち迷走神経➔肝経路を介して肝免疫抑制様機能を維持していることが示唆される(下)。(B)CMT試験における総体計測値、(左-右)総体重、肝臓重量、肝臓:体比。(C)フローサイトメトリーによるイムノプロファイリング。(D)Cのフローサイトメトリーデータの細胞数を組織重量で正規化したもの。(左-右)CD8+ T細胞(CD3+CD8+)、NKT細胞(CD3+CD1d tetramer+)、マクロファージ([non-MDSC] CD3-CD11b+F4/80+)、およびmMDSC集団(CD3-CD11b+F4/80+Ly6C+Ly6G-)。(E) Expanded Methodologyで報告したPMA/イオノマイシン刺激後の生体外細胞内サイトカイン染色。(上)CD4+細胞および(F)NKT+細胞におけるサイトカインマーカーのMFIおよび(下)親の頻度。(G)CD8+マーカーの親度数とMFI、IL10+染色を報告。(H) 近位結腸(10cm)からのRT-qPCRデータ(左-右)Cldn2、Cldn4、Tjp1、Tnfα発現。データはGapdhで正規化し、SVNT相対発現からの倍数変化を報告した。統計的有意性は一元配置分散分析(one-way ANOVA)とTukeyの多重比較(post-hoc)により決定した。棒グラフは平均値±SEMを表す:*P < 0.05; **P < 0.01; ***P < 0.05; ****P < 0.001; ns =有意ではない。
CMTレシピエントは同程度の体重と肝臓重量を示した(図s14B)。フローサイトメトリーによる免疫プロファイリングでは、肝免疫細胞組成の変化が同定され、特にTレシピエント(SVT、HVT)ではSVNT、HVNTレシピエントと比較して肝mMDSC細胞が減少していた。対照的に、CD8+ T細胞の蓄積はCMT群間でほぼ同等であった(図s14C、D)。次に、PMA/イオノマイシン刺激肝リンパ球における細胞内サイトカイン発現を評価した。Live+CD8+T細胞を部分的に教師付きクラスタリングしたところ、MFI発現に基づいて、サイトカインとサイトカイン陰性(Cyto-)のサブセットが明瞭になった(図5H)。CD8+T細胞のUMAPをグループ別に解析した結果、SVNT、HVNT、SVTの各プロファイルで同等のランドスケープが観察された。HVTのUMAPは、Cytoサブセットの相対的減少とc4_CD8_IFNγ+およびc5_CD8_GB+クラスターの拡大によって特徴付けられた(図5H)。フローサイトメトリーデータセットの手動ゲーティングにより、肝リンパ球全体、特にCD8+ T細胞内での細胞傷害性発現(グランザイムB+)および炎症性発現(IFNγ+TNFα+)の上昇が明らかになった(図5I、Jおよび図s14E-G)。
要約すると、肝細胞癌ドナー(担癌マウス)からの微生物叢移植は糞便微生物叢を変化させ、HVTレシピエント内では肝抗腫瘍活性を引き出す。SVTマウスもHVTマウスも同一のドナーから糞便移植を受けたが、HVT肝臓はCD8+ T細胞エフェクター機能の増加を示し、HV肝癌モデルにおける抗腫瘍反応と同等であった(図3)。これらの所見は、腸-肝臓の直接的な活性化を否定するものではないが、上皮組織のRT-qPCRから、様々なクローディン遺伝子やタイトジャンクション遺伝子(Cldn2、Cldn4、Tjp1)を含む、同等の炎症マーカーやバリアマーカーが検出されたことから、HVTの反応は、CMTによるリーキーガットおよび/または腸の炎症によって引き起こされるものではないことが示唆された(図s14H)。
我々は、コリン作動性迷走神経障害が抗腫瘍免疫を促進し、腸内細菌叢によって形成される迷走神経-肝臓アークを介して、肝臓がんにおけるCD8+ T細胞の細胞傷害性を促進することを報告した。腸内細菌叢は肝免疫の組成、機能、免疫抑制に影響を及ぼすが(33, 60-63)、現在進行中の研究では、主に腸内細菌叢と肝臓の直接的な相互作用が研究されている。われわれが得た知見を総合すると、特定の細菌が迷走神経活性を介して肝耐性を維持していることが示唆される。
肝癌は広範な腸内細菌異常症を促進する(64)。興味深いことに、T細胞レシピエントは、以前消化管迷走神経活性化を促進することが報告された硫酸還元細菌の相対量が増加していた(図5Gおよび図s13F)(17, 51)。コリン作動性迷走神経活性は肝臓の免疫寛容を維持し、炎症性T細胞および細胞傷害性T細胞の反応を抑制する。迷走神経活動の下流での破壊(例えば肝迷走神経切断術による)は、ムスカリン性AChRシグナルを介して抗腫瘍免疫を促進し、特にCD8+ T細胞のエフェクター応答を促進する。この研究により、肝性抗腫瘍を抑制するコリン作動性アークが明らかになった(図5K)。今後の研究により、肝がん治療をサポートする微生物および神経免疫の治療標的が明らかになることが期待される。
考察
肝臓がんは最も死亡率の高いがんのひとつである(1, 2)。臨床的進歩にもかかわらず、確立された免疫療法は免疫応答が乏しいため、患者のほとんどが失敗している(9, 65)。外科的切除や切除術が肝癌の唯一の根治的治療法であることに変わりはない(65)。肝抗腫瘍免疫を促進するためには、癌神経科学的取り組みを含む新しいアプローチが必要である。
臓器間のクロストークは免疫応答を制御する(15)。肝悪性腫瘍において、臓器間の発見は主に腸-肝軸に焦点を当てている(9, 66)。実際、腸内細菌叢の調節は、代謝性肝疾患における有望な治療法を明らかにした(66-69)。
がんにおいては、腸内細菌と微生物代謝産物が診断バイオマーカーとして最初に評価された(62, 63, 70)。腸内細菌叢の異常は肝臓がん全体で広く報告されており、特定の分類群と進行ステージとの相関が認められている(71, 72)。画期的な研究により、腸内細菌叢が炎症調節を介して黒色腫がんの転帰と免疫療法の有効性を変化させることが明らかになった(73, 74, 61)。肝細胞癌モデルでは、抗生物質への曝露がNKT細胞の蓄積を増加させ、マウスの肝腫瘍を抑制した(33, 57)。これらの所見から、肝臓の抗腫瘍免疫応答の動的な臓器間調節が明らかになった。
我々は、腸-脳シグナル伝達がACh活性を介して肝免疫寛容に寄与していることを提唱する。正確な肝迷走神経切断は、コリン作動性免疫相互作用を破壊し、腫瘍増殖を抑制する。HVマウスは、NKTの拡大やMDSCの制御を含む広範な免疫変化を示したが(図3A)、Rag1KO研究とCD8+ T細胞枯渇研究(図2)では、CD8+ T細胞反応に重要な役割があることが強調された。健康な肝臓における迷走神経支配の程度はまだ不明であるが(75)、コリン作動性神経線維が肝悪性腫瘍や臨床肝癌で報告されている(26, 76, 77)。コリン作動性シグナル伝達、特にCHRM3を介したシグナル伝達は、転写経路の調節を介して胃癌の増殖に関連していることが以前に報告されている(21, 45)。これらの結果を総合すると、AChはCD8+ T細胞のエフェクター機能を阻害し、CHRM3に依存したACh-CD8+ T細胞軸を同定したことになる。
最近、Zhengら、2023(25)は、CD4+ T細胞におけるコリン作動性シグナル伝達の内在的喪失(Chatノックアウト)が肝がんを悪化させ、Tregの拡大を促進することを報告した。対照的に、迷走神経切断術はTregの蓄積に影響を及ぼさず(図s3B)、肝腫瘍負荷を制御した(図1D)。我々は、ACh活性がCD8+T細胞においてサブセット特異的な応答を示すことを示したが、これはおそらく癌の転帰の違いを説明するものであろう。ACh活性の迷走神経障害によりエフェクターCD8+応答が維持される一方(図2E)、CD4+ T細胞内の遺伝子Chat切除により、オートおよびパラクリンAChシグナル伝達が阻害され、T細胞の機能不全が誘発され、その後肝腫瘍の進行が促進された(25)。
腫瘍の軌跡を超えて、迷走神経の障害は、腫瘍依存性の微生物叢異常と関連する神経症状である疲労を改善した。実際、肝細胞癌マウスの微生物叢は、迷走神経切断マウスでのみ抗腫瘍免疫機能を引き起こすのに十分であった(図5K)。肝細胞癌ドナーからの微生物叢移植は腸内細菌叢プロフィールを変化させ、特に硫化水素産生分類群(すなわち、Desulfovibrio、Bilophila)は大腸癌や肝臓癌と関連している(78, 79)。最近、微生物由来の硫化水素がカルシウム選択的迷走神経受容体の調節を介して迷走神経求心性神経を活性化することが報告され(58)、肝細胞がんが迷走神経-脳シグナルを誘導するメカニズムの可能性が強調された。腸内細菌叢の変化は迷走神経活動に影響を及ぼすが(17)、腸内細菌叢が迷走神経求心性神経を刺激する正確な経路についてはまだよくわかっておらず、今後の研究が必要である。迷走神経求心性神経は腸管上皮を通過しないため、微生物の調節は、特定の細菌に関連するのではなく、複雑なシグナル伝達機構を介して起こる可能性が高い。考えられるメカニズムとしては、迷走神経による小腸内細菌叢の変化、腸内細菌の代謝産物、細菌によって合成される神経伝達物質、微生物によって誘発される炎症、および/または迷走神経-腸内分泌細胞相互作用の細菌による調節などがある(53, 80)。肝枝迷走神経切断術は、総肝枝から分岐する胃十二指腸線維の消失により、上部(十二指腸)腸管の神経支配を部分的に低下させる。実際、迷走神経切断術が小腸の微生物叢に、大腸や糞便の微生物叢よりも強い影響を及ぼす理由は、肝迷走神経解剖学的構造にあると考えられる。
最後に、腸内細菌叢と脳のシグナル伝達が迷走神経活動を介して肝臓の免疫寛容を維持することを示した。これらの知見を総合すると、ACh-CD8+ T細胞軸が標的となりうることが明らかになり、ダイナミックな迷走神経-肝シグナル伝達が同定された。微生物-神経免疫クロストークを利用することで、免疫療法の有効性を向上させる治療戦略が得られ、個別化医療や肝癌の治療成績に示唆を与える。
方法の概要
詳細な方法論と研究材料は、補足テキスト(Expanded Methodology)に記載されている。簡単に説明すると、外科的迷走神経切断術を行い、神経が左横隔膜下迷走神経幹から分岐するところで肝迷走神経枝を剪断するか、あるいは迷走神経の完全性を維持するために偽手術を行った(29)。回復後、一部のマウスにRIL175、B16-F10、A20癌細胞株の肝内注射または尾静脈注射を行った(32-34, 81)。私たちの研究室では、Greten研究室内で開発された免疫パネルを利用し、NCI Flow Cytometry Core and Single-Cell Analysis Facilityと共同で、肝臓、腫瘍、脾臓の免疫サンプルからハイスループットのフローサイトメトリーまたはscRNA-seqを行った。ダウンストリーム解析と可視化は、FlowJo(vs 10+)、Partek Flow、RStudio(R vs 4.2.1+)、Prism(vs 9+)で行った。実験デザイン、バイオインフォマティクス解析、およびデータの利用可能性については、Expanded Methodologyにさらに詳述されている。
リンパ球コリン作動性活性の実験的評価は、フローサイトメトリー、ProcartaPlexイムノアッセイ(Luminex®)、ELISA、RT-qPCR、MTTアッセイ、および/またはACh、ベタネコール、4-DAMPを用いた細胞毒性/細胞内サイトカインアッセイによって行われた。試験管内で得られた知見は、報告された方法論に基づくベタネコール-腫瘍モデルにおけるin vivo研究(22)、(34)によって導かれたCD8+ T細胞枯渇研究、およびJürgen Wess(NIH-国立糖尿病消化器腎臓病研究所[NIDDK])の好意により提供されたChrm3f/fマウス交配ペアを用いたCre/lox交配戦略によって裏付けられた。Expanded Methodologyに記載されているin vitro研究および動物腫瘍モデルのための徹底した実験方法論。
システム生物学的研究では、神経学的研究とマイクロバイオーム研究を行った。行動学的試験には72時間フェノタイピング(PhenoTyper、Noldus)、Y迷路、オープンフィールド試験が含まれ、NIH-NHLBI Murine Phenotyping Coreにより実施された。病理組織学的検査はFrederick National LaboratoryのMolecular Histopathology Laboratoryで行われ、標準的なH&E染色(肝臓)と脳幹のcFOS神経細胞染色が含まれた。微生物叢ドナーは、RIL175細胞または非腫瘍ビヒクルコントロールの肝内注射を受けたマウスであった(糞便内容物、腫瘍発生21日後)。ポリエチレングリコール(PEG;下剤)処理後に微生物叢をレシピエントに移植し、先行移植研究(57, 82)に基づいた方法で、糞便内容物を濾過し、50%グリセロールで保存した。NIH Laboratory of Integrative Cancer Immunology-Microbiome and Genetics Coreにおいて、16S rRNA-Seq用の糞便サンプル、糞便、小腸(空腸/回腸)内容物を収集し、ライブラリー調製と配列決定を行った。サンプルはQIIME2(83)およびLefSE(84)パイプラインで解析した。詳細はExpanded Methodologyに記載されている。
さらに著者らは、MedStar Georgetown University Hospital(Washington DC、IRB #2017 -0365)で実施された臨床HCC手術検体から最近接細胞解析を行った。手術後、細胞プロファイリングはco-detection by indexing(CODEX)イメージングを介して実施され、National Cancer Institute Halo Image Analysis Resourceを介してアクセスされた。方法論とサンプル処理は(36)に報告されている。パスウェイ濃縮解析(ReactomePA)は、バルクRNA-Seqデータセットと(35)で提供された差次的発現遺伝子リストから実施した。残りの方法論、材料、解析、データ保存はExpanded Methodologyに記載。
資金提供
KCBは、NIHからのiCURE-NCIフェローシップ(2020年)、Sallie Rosen Kaplan Women in Science Funding(2023年)、SITC-Genentech Women in Science Fellowship(2023年)による資金援助を受けている。B.R.は現在、ドイツのエクセレンス戦略(EXC 2180 - 390900677)の下、ドイツ研究振興財団(DFG、ドイツ研究財団)から研究助成を受けている。FJR-MはNIH医学研究奨学生プログラムによる資金援助を受けている。TFGはNIH、NCIのIntramural Research Program(ZIA BC011345, ZO1 BC010870)から研究助成を受けた。TFGとGTはNCI FLEX awardの支援を受けた。
著者貢献
KCBは、Greten、Trinchieri、Xieの各研究室のメンバーとの共同研究により、プロジェクトのコンセプト立案、手順の最適化、実験の実施、データ解析/可視化の準備を行った。原稿は共著者全員の意見を参考にKCBが執筆した。RT、BR、YM、M-RB、CM、MD、AN、JQ、PH、MS、BLG、SW、DAS、FJR-M、JM、SG、SNG、BD、JG、RR、AKDは、手順の設計、実験の実施、および/または解析を行った。CX、GT、FK、TFGは研究室メンバー、プロジェクト開発、リソース、分析解釈を指導した。TFGは資金獲得、研究チーム間の共同監督、科学的仮説の概念化、研究および論文作成の指導、最終原稿の編集を主導した。
競合利益
著者らは、競合する利益はないと宣言している。
拡張された方法論
マウスおよび迷走神経切断術
すべての動物手順および実験エンドポイントは、National Research Council's Guide for the Care and Use of Laboratory Animalsに従ってNCI Institutional Animal Care and Use Committeeの承認を得た。マウスの処置は動物実験計画書MOB-028、TGOB-013、TGOB-015に記載されている。マウスはCRC動物施設およびNHLBI Murine Phenotyping Coreで12時間の明暗サイクルで飼育し、餌と水を自由に摂取させた。
雌雄マウスはCharles River Laboratoriesから購入した: C57BL/6(系統コード:#556)、BALB/c(系統コード:#028)、C57BL/6-Tg(TcraTcrb)1100Mjb/J(OT-1;系統コード:#003831)、またはJackson Laboratoriesから購入した: C57BL/6J(系統コード:#000664)、C57BL/6-Tg(Cd8a-cre)1Itan/J(CD8cre系統コード:#008766)、C57BL/6J-Rag1<em10lutzy>/J(Rag1KO系統コード:#034159)。MR3-floxマウスは、Yamadaら、2001(85)に記載されているように作製され、Jürgen Wess博士(NIH-NIDDK、Bethesda、MD 20892)の好意により提供された。CD8creM3Rf/fマウスを開発するために、CD8creマウスをMR3-flox動物と交配し、CD8creMR3WT/f子孫をMR3-flox動物と戻し交配した。同腹仔コントロールにはCD8cre、M3Rf/f、CD8creMR3WT/fが含まれた。繁殖はCRC動物施設内の研究者が行い、遺伝子型判定はTransnetYXサービスを通じて確認した。処置は、2~4ヵ月齢の雌雄を一致させたコントロールと、8~12週齢のマウスに行った肝迷走神経切断または偽迷走神経切断を利用した。
肝迷走神経切断または偽外科的コントロールはCharles River Laboratoriesから入手した(外科的サービスは2022年に終了)。あるいは、Terataniら、2020(29)に記載されているように迷走神経切開を行った。簡単に説明すると、剣状突起から腹部中央まで垂直に切開した。外科用ルーペ(4.0X、作動距離340mm)を用いて、横隔膜から下降する左(前)迷走神経枝から総肝枝を同定した。肝枝を分離し、外科用ピンセットで切り取った。偽手術では、肝枝は同定されたが切断されなかった。癒着を防ぐため、腹腔内を滅菌した0.9%生理食塩水で満たした。筋切開は外科用縫合糸で閉鎖し、皮膚フラップは創傷クリップで閉鎖し、鎮痛剤で経過観察した。手術と手術の間には2週間の回復期間を設けた。
細胞株と腫瘍モデル
Kapanadzeら、2013(86)に記載されているように、マウス肝細胞がん細胞株RIL175をRPMI1640培地(Thermo Fisher Scientific、カタログ番号61870127)+10%FCS(糞便コス血清:Corning Inc.、カタログ番号MT35010CV)で培養した。B16-F10(マウスメラノーマ)細胞株は、Kreiterら、2015(87)に記載されているように、Ugur Sahin博士(ドイツ、マインツ)から提供され、A20(マウスリンパ腫)およびB16-F10-OVA細胞株はATCCから購入した。in vitro研究および転移性腫瘍細胞株については、細胞培地(RPMI1640+10%FCS)にさらに、1mMピルビン酸ナトリウム(Thermo Fisher Scientific、カタログ番号11360070)、1mM-1M HEPES(Thermo Fisher Scientific、カタログ番号15630080)+1mM NEAA(Thermo Fisher Scientific、カタログ番号11140050)および/または500ng/mLピューロマイシン(InvivoGen)、および/または3. 5Mβ-メルカプトエタノール(Sigma-Aldrichカタログ番号60-24-2)。ペニシリン-ストレプトマイシン溶液1%(Thermo Fisher Scientific、カタログ番号5070063)は、細菌汚染を防ぐために添加した。アリコートは使用前に12回未満継代し、Rufら, 2023 (36)に記載されているプロトコールを参照した。細胞株は、ルーチンの細菌およびマイコプラズマ検査(2022年5月、Molecular Testing of Biological Materials NCI, Frederick 21702)に従って汚染陰性であった。
腫瘍細胞の開始は、Chiら、2018(33)に記載され、Brownら、2018(81)によって報告された方法論に詳述されているように行った。肝内モデルについては、2.5×105個の癌細胞とマトリゲル(Corning Inc.、カタログ番号354230)を含む1:1滅菌PBSからなる20μL/マウス注射を、肝臓の左側葉に注入した。硝酸銀(ベットワン社、カタログ番号NDC13895-706-10)で肝出血を止め、切開部を滅菌縫合糸と創傷クリップで閉鎖した。皮下(骨盤)注射と尾静脈注射には、滅菌PBSに1.0×106個の癌細胞を100μL/マウスを入れた。マウスは手術前に無作為に割り付けられた。ルシフェラーゼ発現RIL-175細胞のin vivoイメージング(IVIS Spectrum®)は、Xenogen IVIS 100 Imagingテクノロジーを用いて行った。マウスは150mg/kgのルシフェリン(REGIS Technologies、カタログ番号2591-17-5)を腹腔内注射した7.5分後にイメージングを受けた。特に断りのない限り、腫瘍重量は投与開始後21日目に測定した。マウスは、実験終了前(すなわち、外科的処置中)、または不適切な外科的開始(すなわち、肝臓以外の腹部臓器への偶発的注射)により死亡した場合のみ、解析から除外した。
アセチルコリン作動薬モデル
ムスカリン治療は、Renzら、2018(22)で報告されたプロトコールに従った。塩化ベタネコール(Millipore Sigma Aldrichカタログ番号1071009)を飲料水中に投与した(400μg/mL)。アゴニストへの曝露は腫瘍形成の3-5日前から開始し、実験手順を通じて新鮮な水を1-2X/週補充した。マウスは飲料水を自由に摂取できた。ベタネコール投与マウスと水のみの対照マウスでは、同程度の飲水量が観察された。
CD8 T細胞の枯渇
CD8+T細胞の枯渇は、200μg/マウスの抗体(InVivoMAb anti-mouse CD8α、Clone 2.43、BioxCell)を含む100μLの腹腔内注射によって行われた。マウスには腫瘍発生1日前に注射し、その後実験終了まで7日ごとに注射した。アイソタイプ適合IgG2b(InVivoMAbラットIgG2bアイソタイプコントロール、抗キーホールリンペットヘモシアニン、クローンLTF-2、BioxCell)を、IgGコントロールに200ug/マウスで投与した。
微生物叢移植
年齢をマッチさせたRIL175腫瘍マウスおよび非腫瘍コントロールマウス(腫瘍発生後21日)のセカルパウチを微生物叢移植に利用した。個々の糞便内容物を滅菌した50%グリセロール溶液(5 mL)に添加した。混合後、糞便内容物を100μmのフィルターで濾過し、1mLずつ-80℃で保存した。レシピエントマウスをPEG(ポリエチレングリコール、Millipore Sigmaカタログ#25322-68-3)で処理し、Wrzosekら、2018(82)によって報告されたように、セカル16S DNAコピーを90%減少させるプロトコルを利用して腸内マイクロバイオームを枯渇させた。簡単に述べると、マウスは20分ごとに4回の経口ガベージ(滅菌蒸留H2O中の425g/L PEGを含む200μL)を受けた。最後のPEG処理後、マウスは最初のCMTを経口投与(100μL)された:d 1。経口投与は3、5、8日目に繰り返した。16SrRNA-Seqによる微生物叢移植の成功を確認するため、実験前および実験期間中、糞便ペレットを採取した。すべての実験は最終経口摂取から10日以内に終了した。
行動および活動表現型解析
表現型解析を行ったマウスは、NHLBI(国立心肺血液研究所)内のMurine Phenotyping Coreで12時間の明暗サイクルで飼育した。すべての動物処置はNCI Division of Intramural Research Animal Care and Use Committee(提案TGOB-015)に従って行われた。記録試験の前に、マウスは馴化と穏やかな取り扱い期間を経た。解析は、迷走神経切開手技についてはベースライン(迷走神経切開後)、早期(肝内注入後7日未満)、後期(肝内注入後7日以上)、およびベースライン(cecal microbiota移植前7日未満)およびCMT(初回細菌経口投与7日以上)で繰り返した。
フェノタイパーおよびXYケージモニタリング
マウスは、赤外線カメラトップユニットを装備したPhenoTyperホームケージ(Noldus社、オランダ、ワーヘニンゲン)でホームケージモニタリング(72時間/ラウンド)を受けた。PhenoTyperケージからの映像は、EthoVision XT行動追跡ソフトウェアを用いて解析し、マウスの活動と動きを測定した。ホームケージモニタリング中、マウスは単独飼育され、ALPHA-driの寝具が与えられ、餌と水が自由に摂取できた。
オープンフィールドテスト
マウスをホームケージから取り出し、16'×16'×16'Perspex®アリーナのビューイングチャンバーに個別に入れ、自由に探索させた。マウスはホームケージに戻る前に30分間記録された。距離、速度、移動度、定義されたゾーンでの滞在時間はANY-maze行動追跡ソフトウェア(Stoelting Co, Il)を用いて分析した。
自発Y字迷路
スタートアーム(18.4cmのスタートボックスと30.5cmの走路がドアでつながっており、中央のハブから2本の36.8cmのサイドアームが互いに120°の角度で離れている)からなる3本腕のポリカーボネート製Y字迷路にマウスを入れた。マウスをスタートボックスに入れ、ドアを開け、5分間迷路を自由に探索させた。アームの総訪問回数をスコア化し、交替率(総交替回数/可能な3本鎖×100)を手動でスコア化した。
臨床解析 Kaplan-Meier プロット
Kaplan-Meierプロットは、Kaplan-Meierプロッター機能(88)およびHuman Protein Atlas(89)(2023年12月にアクセスされたウェブサイト)を介してTIMER2.0上でCHRM3を評価することにより決定された。
リアクトームPAとRNA-Seq
がんにおけるコリン作動性および神経経路の変化を調べるために、Aranら、2017(35)で提供されたデータセットを利用した。ここで著者らは、健常組織と腫瘍関連組織を評価する8つの腫瘍型にわたる6506の生検について、遺伝子発現の差分解析を行った。ReactomePAパッケージ(90)を使用し、RStudio(バージョン2022.07.1)で作成した可視化により、公開された補足データ1、健常組織と腫瘍隣接(NAT)組織間の差分発現解析についてReactomePA解析を行った。
CODEX解析
37プレックスのCODEXパネルを用いて、肝細胞癌(HCC)患者を対象にCODEX(Co-detection by indexing)多重化組織イメージングを行った。MedStar Georgetown University Hospital(Washington DC)での募集プロセスにおいて、性別と民族の多様性を確保する努力がなされた。生検は外科的HCC切除を受けた患者から採取した。臨床データおよび生物学的データの収集に関するインフォームド・コンセントとデータの適切な移転/保存は、Institutional Review Boardのプロトコル#2017-0365およびMaterial Transfer Agreement #43655 -18に従った。CODEXの生画像はThe Cancer Imaging Archives (TCIA)でhttps://doi.org/10.7937/bh0r-y074。臨床的および病理学的情報、検体の調製、抗体の検証、およびCODEX多重免疫蛍光法および画像化、ならびにデータの所在は、Rufら、2023(36)に提供されている。
最近接神経解析のために、NCI HALO Image Analysis Resourceから12個のHCC標本の高解像度一次画像をレビューした。神経軸索は、非核(DAPI陰性)、非胆管(panサイトケラチン陰性)、NCAM+枝(神経細胞接着分子+、50μm以上)に分類した。各標本で3つの神経リッチ領域が同定された。各領域で異なる抗体マーカーを用いた同じサイズの画像(100倍ズーム)を撮影し、最も近い細胞近傍を手動で同定した。画像1(構造/間質パネル)、画像2(リンパ系パネル)、画像3(骨髄系パネル)。その後、画像(jpg、tifファイル)をImageJ(FIJI)で解析し、最近接細胞を手動で決定した。構造/間質細胞、骨髄系細胞、リンパ系細胞を同定するために、選択した数のタンパク質マーカーを用いた(補足表2参照)。
免疫細胞の単離とフローサイトメトリー
脾臓、肝臓、および腫瘍浸潤細胞の単細胞分離は、報告されているプロトコル、すなわちHeinrichら、2021(91)およびRufら、2023(36)に従った。安楽死後、動物から臓器を摘出し、固形腫瘍(RIL175、B16-F10)を肝臓から摘出するか、腫瘍性肝臓(A20)として処理した。脾臓と肝臓の両方をホモジナイズし、70μmのナイロンメッシュで濾過し、4℃で15分間遠心分離した(400倍RCF)。肝臓はさらに90%パーコール(Cytiva、カタログ番号17089101)による密度勾配遠心分離を行い、間質成分から免疫細胞を分離した。ホモジナイズ後、gentleMACS Octo Dissociator(Miltenyi Biotecカタログ#130-096-427、マウス腫瘍プログラム)を用いて腫瘍を解離し、その後Lympholyte Cell Separation Media(Cedarlane Laboratories、カタログ#CL5035)を用いて密度勾配遠心した。全てのサンプルをACK-Lysis Buffer(Quality Biologicals、カタログ番号118-156-721)で処理し、赤血球を除去した。単離した単細胞を70μmのナイロンメッシュに2回通し、残存する残屑を除去し、染色前にサンプルをPBS+10%FCS中-4℃で保存した。
染色はすべて4℃で行った。マウスサンプルは以下のように染色した: Zombie UV(TM) Fixable Viability(BioLegend、カタログ番号423108)を用いて、PBS中で20分間Live/Dead染色を行い、その後FACSバッファー中で10分間Fc-blocking(eBiosciences、カタログ番号14-0161-85)を行った。FACSバッファーには、9.7%10X PBS(コーニング社、カタログ番号46-013-CM)、20% FCS、0.1%アジ化ナトリウム(シグマアルドリッチ社、カタログ番号26628-22-8)を用いた。表面染色は30分間行い、抗体は次のものを含む:Alexa Fluor 700上の抗CD45(BioLegend、カタログ#103128、クローン#30-F11)、Alexa Fluor 700上の抗Ly6G(BD Biosciences、カタログ#561236、クローン#1A8)、BV605上の抗CD4(BioLegend、カタログ#100547、クローン#RM4-5)、APC/Cy7上の抗Ly6C(BioLegend、カタログ#128026、クローン#HK1. 4)、PE上の抗CD3(BioLegend、カタログ#100206、クローン#17A2)、PerCp-Cy5上の抗CD19(BioLegend、カタログ#128026、クローン#HK1. 5(BioLegend、カタログ#152406、クローン#1D3/CD19)、FITC上の抗F4/80(BioLegend、カタログ#123108、クローン#BM8)、パシフィックブルー上の抗CD11b(Biolegend、 カタログ番号101224、クローン番号M1/70)、PE/Cy7上の抗TCRβ(BioLegend、カタログ番号109222、クローン番号H57-597)、BV510上の抗MHCII(BioLegend、カタログ番号107635、クローン番号M5/114. 15.2)、BV650の抗CD11c(BioLegend、カタログ番号117339、クローン番号N418)、BV785の抗CD8(BioLegend、カタログ番号100749、クローン番号53-6. 7)、Alexa Fluor 700の抗CD45R/B220(BioLegendカタログ#103232、クローン#RA3-6B2)、Alexa Fluorの抗F4/80(BioLegendカタログ#123130、クローン#BM8)、Alexa Fluorの抗CD11b(BioLegendカタログ#101222、 クローン#M1/70)、抗CD3はAlexa Fluor 594(Biolegendカタログ#100240、クローン#17A2)、抗TCRβはAPC/FireTM 750(Biolegendカタログ#109246、クローン#H57-597)、抗NK1. 1(BDバイオサイエンス、カタログ番号741926、クローン番号PK136[RUO])、FITC上の抗PD1(バイオレジェンド、カタログ番号135214、クローン番号29F. A12)、およびAPC上のCd1d四量体(エモリー大学との共同研究によるNIH四量体コア施設)を用いた。
細胞内サイトカイン染色に先立ち、細胞はPMA/イオノマイシン刺激を受けた。具体的には、0.4 µg/mL IL2および/またはLeukocyte Activation Cocktail, with BD GolgiPlug™ (BD Biosciences catalogue #550583 )を含む培地(20 µL/mL培地)で4時間刺激した。細胞内染色に先立ち、BD Cytofix/Cytoperm™ Fixation/Permeabilization Kit(BD Biosciences、カタログ番号554714)またはBD Pharmingen™ Transcription Factor Buffer Set(BD Biosciences、カタログ番号562574[FoxP3染色用])を用いて、メーカーの推奨に従って細胞を固定した。その後の細胞内サイトカイン染色(30分)では、以下の抗体を用いた:APC上の抗パーフォリン(BioLegend社、カタログ番号154304、クローン番号S16009A)、APC上の抗IL6(BioLegend社、カタログ番号504508、クローン番号MP5-20F3)、PerCP-Cy5上の抗TNFα(BioLegend社、カタログ番号504508、クローン番号MP5-20F3)。 5(BioLegend、カタログ番号506322、クローン番号MP6-XT22)、抗グランザイムBをFITC(BioLegend、カタログ番号515403、クローン番号GB11)、抗IFNγをPE/Cy7(BioLegend、カタログ番号505826、クローン番号XMG1. 2)、BV421上の抗IL10(Biolegend、カタログ番号505022、クローン番号JES5-16E3)、染色後、サンプルをCytoFLEX LXフローサイトメーター(Beckman Coulter、RRID:SCR_0129627)で評価した。その後の解析は、FlowJoソフトウェア(vs 10、RRID:SCR_008520)またはPartek Flow(NIH HPC Biowulf)を用いて行った。頻度(%生細胞)または細胞数(絶対細胞数:集団頻度 x 総生細胞数 / サンプル重量)が提供された。免疫細胞の絶対数は、頻度に総生細胞数を乗じて算出し、肝臓重量で割った。
scRNA-Seq
セルソーティング、ライブラリー調製、10Xゲノミクスシーケンス
scRNA-Seq用に8サンプルを準備した。固形RIL175腫瘍を腫瘍発生から21日後にSVおよびHV肝臓(n = 3 /群)から摘出した。RIL175腫瘍保有マウス(n = SVマウス3匹、HVマウス3匹)の非腫瘍性肝臓とマッチした(プールされた)腫瘍から。腫瘍および肝臓からの免疫細胞は、以前の方法論で説明したように単離した。表面染色後、Aurora CS Cell Sorter(Cytek® Biosciences、RRID:SCR_XXXXXX)を用いて、NIH Flow Cytometry CoreでLive+CD45+細胞をPBS+0.04%BSA(ThermoFisher Scientific、カタログ番号B14)にソーティングした。その後の細胞数と生存率は、Luna FL Automated Fluorescence Cell Counter(Logos Biosystem)を用いたヨウ化プロピジウム/アクリジン染色で決定した。
サンプルはSCAF(NIH Single Cell Analysis Facility)に提供され、ライブラリー調製、配列決定、データ処理が行われた。細胞数はLunaFx7蛍光セルカウンター(Logos Biosystem)で測定し、6,000細胞/レーンを目標に調整した。ライブラリーの調製とサンプルのロードはSCAFで行い、10X Genomics Single Cell User Guideに従い、Chromium Connect(10X Genomicsオートメーションプラットフォーム)を使用した。
シーケンシングはNovaSeq 6000 Reagent Kit(Illumina)を用いて行った。5' Immune Profiling v2 Dual-Index chemistry(10x Genomics)を用いて2回のシーケンスラン(NovaSeq 6k S1 100-cycle)を行った。
scRNA-Seq処理
データのベースコールおよびクオリティスコアリングにはRTA 3.4.4(イルミナ)を使用した。その後のデータ処理はCell Ranger v7.1.0(Illumina)で行い、マウスゲノミクスリファレンス(refdata-gex-mm10-2020-A)へのデマルチプレックス(Bcl2fastq 2.20.0)およびアライメント(STAR 2.7.2a)を含む。処理後、Mean Reads per Cell = 83,551、Total Genes Detected = 19,745、Median UMI Counters per Cell = 6,237(全サンプル出力の平均値)。配列の飽和度は81%~93%であり、配列データのマウストランスクリプトームへの頑健なマッピングが示された。
scRNA-Seq解析
データはBiowulf(NIH HPC Linux cluster)に置かれ、RStudio(vs 2023.09.1+494)下のSeuratパッケージ(vs 4.3.0.1)を用いてダウンストリーム解析が行われた。カウントマトリックスファイルはRead10X_h5関数を用いて読み込まれ、10細胞未満の発現遺伝子を除いたSeuratオブジェクトに変換された。低品質細胞は以下の基準で除去した:featuresが500未満または4000遺伝子以上の細胞、nCountsが500未満または14000以上の細胞、log10(features/counts)<0.8、ミトコンドリアRNA含量が20%以上の細胞。各サンプルのダブレットはDoubletFinder(vs 2.0.3)を用いて除去した。正規化は個々のサンプルでSCTransform関数を用いて行い、その後すべてのサンプルを1つのSeuratオブジェクトにマージした。バッチ補正にはHarmony(vs 0.1.1)を適用した。
FindClusters関数を用いて解像度0.4で適切なクラスターを見つけ、T細胞、B細胞、NK細胞、ミエロイド細胞の個別のクラスターを作成した。T細胞のさらなるサブクラスター化は、分解能0.45でFindClusters関数を用いて行った。遺伝子差解析は、FindAllMarkers()を用いて全てのクラスターについて、また、FindMarkers()を用いて特定のメモリークラスターを分離するために、min.pctを0.1,logfc.thresholdを0.1,only.posをTRUEとして、複数の解像度に渡って独立として持続するクラスターについて行った。UMAPはRunUMAP関数を用いて作成した。遺伝子発現の差はEnhanced Volcanoパッケージ(バージョン1.20.0)を用いてプロットした。
AChR+およびAChR-判定
すべてのアセチルコリン受容体遺伝子(Chrm1, Chrm2, Chrm3, Chrm4, Chrm5, Chrna1, Chrna2, Chrna3, Chrna4, Chrna5, Chrna6, Chrna7, Chrna8, Chrna9, Chrna, Chrnb1, Chrnb2, Chrnb3, Chrnb4, Chrnd, Chrne, Chrng)の発現に基づき、AchR判定のためのscGSEAを完成させた。遺伝子発現の二峰性のバイオリンプロットから、適切な閾値を0.05とした。
パスウェイ濃縮解析
マウスデータベース(org.Mm.eg.db)を用いて、clusterProfiler(version 4.10.0)パッケージを肝臓と腫瘍に分けてパスウェイ濃縮解析に使用した。clusterProfilerのカットオフはp値0.05、機能濃縮GO、生物学的プロセス(BP)が選択された。
単クローン軌跡解析
Monocle3パッケージ(バージョン1.3.4)を用いて、Monocole解析とPseudotime解析を行った。Monocleでは、Leidenクラスタリング法を解像度7e-4で選択した。
in vitro AChシグナル伝達アッセイ
細胞内活性化アッセイ
ACK Lysing bufferで処理したマウス脾臓細胞からリンパ球を増殖させた。CD3+ T細胞は、1μg/mlの抗マウスCD3(eBiosciences社、カタログ番号16-0031-82、クローン番号145-2C11)と0.1μg/mlの抗マウスCD28(eBiosciences社、カタログ番号16-0281-85、クローン番号37.51)抗体を介して拡大し、IL2(100ng/mL)またはCD3/CD28 Dynabeads(ThermoFischer Scientific社、カタログ番号11452D)とCD3+ T細胞の3:1の比率で活性化した。48時間培養後、リンパ球は薬剤暴露(2時間)を受け、次いで薬剤暴露による活性化(4時間)を受けた。4時間の活性化には、0.4μg/mLのIL2および/またはLeukocyte Activation Cocktail、BD GolgiPlug™(20μL/mL培地)を用いた。薬理学的化合物としては、塩化ACh(Sigma Aldrich、カタログ番号A2661-25G)、塩化ベタネコール(Sigma Aldrich、カタログ番号071009-200MG)、または4-DAMP(Sigma Aldrich、カタログ番号SML0255-50MG)を用量依存的に用いた。薬理学的濃度は、組織ELISA、マウス(22)およびin vitroの解析結果(92)から決定した。細胞内サイトカインおよび細胞毒性発現レベルは、フローサイトメトリーにより評価した。
細胞毒性アッセイ
B16-F10-OVA細胞を、CellTrace™ Violet Cell Proliferation Kit(Invitrogen、カタログ番号C34557)を用いて製造元の仕様に従って標識し、96ウェルプレート内に1.5×104個/ウェルでプレーティングした。12時間培養後、CD8+ T細胞を報告されたエフェクター対ターゲットの比率で共培養した。これらのCD8+ T細胞は、OT-1脾臓細胞からACh(1μg/ml)を添加または無添加で5日間培養し、CD8a+ T Cell Isolation Kit, mouse(Miltenyi Biotec, カタログ番号130-104-075)を用いてCD8+ T細胞を濃縮した。共培養後、腫瘍細胞を剥離し、回収した上清と合わせた。Zombie UV™ Fixable Viability Kitによる生死染色を行い、生存率を評価した。加熱死滅させた共培養は、最大細胞毒性のコントロールとなった。最大細胞毒性のパーセンテージは、式[(コントロールの生細胞数-実験条件の生細胞数)/(コントロールの生細胞数-熱殺コントロールの生細胞数)]を用いて計算した。
別の同等の共培養細胞は、Brefeldin A(Biolegend、カタログ番号420601)で4時間処理した。CD8+T細胞の細胞内サイトカインおよびエフェクター分子の発現をフローサイトメトリーで評価した。
MTTアッセイ
腫瘍細胞株を、腫瘍細胞株培地(RPMI +10% FCS +1% penicillin-streptomycin溶液)中、96ウェルプレートに10,000細胞/ウェルでプレーティングした。48時間後、培地を除去し、製造業者のガイドラインに従ってMTT細胞増殖キット(Abcam、カタログ番号ab211091)を用いて生存率を測定した。
ELISA法
アセチルコリン含量はQuickDetect™ ACh (Mouse) ELISA Kit(Biovision、カタログ番号E4453)を用いて測定した。マウス組織を採取し、1mLのPBSおよびRoche cOmplete™ EDTAフリープロテアーゼ阻害剤(Millipore Sigma、カタログ番号11873580001)を含む2mL Lysing Matrix Dチューブ(MP Biomedicals、カタログ番号116913050-CF)に入れた。組織をFastPrep®-24(MP Biomedicals社製)で2ラウンド(5.5 m/s、45秒)ホモジナイズした。ホモジネートを4℃で遠心分離し、冷やした上清を回収して-80℃で保存し、分析に使用した。データは、Pierce™ BCA Protein Assay Kits(ThermoFischer Scientific、カタログ番号23225)で測定した組織重量またはタンパク質含量で正規化した。
RNA単離とRT-qPCR
大腸組織(結腸近位部10cm)を摘出し、RNeasy Mini Kit(Qiagen、カタログ番号74106)を用いて組織RNAを単離した。RNA含量と純度はnanodrop(ThermoFisher Scientific、カタログ番号ND-2000)で測定した。調整したRNA含量は、PCR(Bio-Rad、カタログ#1852148)を介してiScript cDNA Synthesis Kit(Bio-Rad, Cat #1708891 )を用いたcDNA調製に利用した。サンプルはRT-qPCRの前に-80℃で保存した。サンプルはRT-qPCRの前にSsoAdvanced Universal SYBR Green Supermix (Bio-Rad, Cat #1725271 )を用いて調製した。すべてのプライマーはBrownら、2015(93)に記載されている。Gadphは内因性ハウスキーピングコントロールを提供し、ddCT計算の正規化に使用した。
cFOS染色
マウスは100%炭酸ガスで空気を置換することにより安楽死させ、5分以内に組織採取のために断頭した。脳はAP領域を含む脳幹レベルで採取した。脳は10%中性緩衝ホルマリンで最低5日間固定した。固定後、脳組織はSakura® Tissue-Tek® VIP™自動プロセッサーで処理し、Sakura® Tissue-Tek® TEC™包埋センターでパラフィンブロックに包埋した。
すべてのブロックを手動のMIRCROM HM 325ミクロトームで切片化し、荷電スライドに載せた。スライドはH&E染色前に80℃のオーブンで1時間乾燥させた。ヘマトキシリン・エオシン(H&E)染色は、Sakura® Tissue-Tek®Prisma™ automated Stainerを用いて行った。スライドを水和し、市販のヘマトキシリン、清澄剤、ブルーイング試薬、エオシン-Yで染色した。退行染色法を用いた。この方法は、組織を意図的に過剰に染色し、その後、過剰な染色を除去するために分化ステップ(清澄剤/ブルーイング試薬)を使用する。スライドはSakura® Tissue-Tek® Glas™ automatic cover slipperを用いてカバースリップさせ、検討前に乾燥させた。H&Eスライドは、AP領域の有無を確認するために病理医1名が評価した。IHC染色は、LeicaBiosystemsのBondRXオートステイナーを用い、5μmの組織切片に以下の条件で行った: Epitope Retrieval 1 (Citrate) 20'、Normal Goat Serum Block、c-Fos (Synaptic Systems #226 308, 1:1000 incubated 30')、Biotinylated Goat anti-Guinea Pig IgG (Vector Labs)、Streptavidin-HRP (Invitrogen #434323 )、Bond Polymer Refine Detection Kit (LeicaBiosystems #DS9800 ) から PostPrimary と Polymer Reagents を除いたもの。陰性コントロールには、一次抗体の代わりに Buffer を使用した。スライドを Bond autostainer から取り出し、段階的なエタノールで脱水し、キシレンで洗浄し、カバースリップした。
16SrRNA シークエンシング
16SrRNA-Seqには以下のサンプルを使用した:糞便ペレット、小腸(小腸遠位部10cmから採取した回腸/空腸内容物)、盲腸(盲腸内容物)、ドナー内容物(50%グリセロール中の凍結細菌スラリー)。ライブラリー調製に先立ち、サンプルはNIHのLaboratory of Integrative Cancer Immunology-Microbiome and Genetics Core(LICI-MGC)のパイプラインに従い、あらかじめ加熱(10分、60℃)したMBL Solution(Qiagen)に添加した。
ライブラリーの調製と配列決定は、LICI-MGC Coreが確立されたプロトコールに従って行った。DNA抽出にはPowerSoil DNA Isolation Kit(Qiagen、カタログ番号47014)を使用した。16S V4領域(515F:5′-GTGCCAGCAGCCGCGGTAA-3′、806R:5′-GGACTACCAGGGTATCTAAT-3′)の細菌増幅は、epMotion 5073(Qiagen)ロボットで行った。16S DNAはNGS MiSeqプラットフォーム(Illumina)でシーケンスし、ペアエンドリード(2×250塩基対)を作成した。
その後、QIIME2 vs. 2023.7(CITE)を用いて、デマルチプレックス、品質管理フィルタリング、アルファ希薄化、系統多様性解析を行った。フォワードリードはQIIME2に取り込まれ、DAD2アルゴリズム(CITE)が品質管理とフィーチャーテーブル構築を行った。サンプリング深度は、alpha rarefactionとQIIME2のtable.qzv出力に基づいて決定した。分類学的分類は、データセット固有の参照リードとSILVA 132 (99% OUT sequence identity)参照分類法(CITE!)で学習したナイーブベイズ分類器を利用した。線形判別分析 Effect Size (LefSe)分析および可視化は、Huttenhower Galaxy server galaxy.biobakery.org上で行った。) 追加のデータ可視化は、RStudio(vs.2022.07.1)とGraphPad Prism(vs.9+)を用いてR(vs.4.2.1)で行った。
生の16SrRNA-Seq .fasqファイルはNCBI Sequence Read Archive (SRA) public database PRJNA105510に寄託した。
統計解析
実験手順のサンプルサイズは、同一の腫瘍モデルを利用した先行研究および先行解析(34, 36, 91)を参考にした。統計学的検定および結果は、原稿および図の凡例に報告されている。解析にはGraphPad Prism vs 9+ (RRID:SCR_002798) およびR (vs. 4.2.1; RStudio vs. 2022.070.01)を用い、統計的有意性はP <0.05とした。
データの入手
現時点では、データのリクエストはTim F. Greten博士(tim.greten{at}nih.gov)までお願いします。
補足ファイルの凡例
補足ファイル1 腫瘍隣接組織は免疫および神経経路に変化を示す
全癌種に対する経路濃縮解析のアウトプット
補足ファイル2 CODEX同定マーカー
最も近い細胞隣接組織を同定するための18色抗体パネル
補足ファイル3 scRNA-Seq差次発現遺伝子リスト
すべての肝臓および腫瘍のscRNA-Seq免疫細胞から得られたDEGを平均対数倍変化値でランク付けしたもの
補足ファイル4 神経免疫遺伝子リスト
神経免疫パネルのKEGG GO値
補足ファイル5 マイクロバイオーム解析
コホージングおよびCMT実験におけるさらなるβおよびα多様性の統計と解析。
謝辞
著者らは、Sophie Wang、Gary Shaw(Greten Laboratory Managers)、Vanessa Catania、Varun Subramanyam、Gauri Garg、Thien Nguyen、Aviva Leah Menashy、およびXiao Binに感謝する。Jay Berzofsky博士とGreten、Berzofsky、Trinchieri、Xie各研究室の同僚には、神経免疫とマイクロバイオームの相互作用について貴重な議論をしていただいた。
Lee Chedester博士(National Institute on Alcohol Abuse and Alcoholism)、Luiz F. Barella博士(元National Institute of Diabetes and Digestive and Kidney Diseases)、およびMichael J. Reilly博士(Charles River Surgery Supervisor)には、肝迷走神経切開のための技術的設備および/または指導を提供していただいた。MR3 KO繁殖ペアはJürgen Wess博士(NIDDK)の好意により提供された。David Kleiner博士は病理学およびCODEX解釈の指導を行った。Research Flow Facility(National Cancer Institute)のWilliam Telford博士が免疫選別(scRNA-Seq)を検討した。Murine Phenotyping Core(NIH-NHLBI)のメンバー、特にAudrey Noguchi、Morteza Peiravi、Heather Pottsに感謝する。Michaelケリー博士とSingle Cell Analysis Facility(NCI-SCAF)のメンバー、特にJatinder Singh、Charlie Seibert、Kimia Dadkhah、Ian Taukulisには、scRNA-Seqの準備と一次データのデポジションについて感謝する(Frederick National Lab契約番号#75N91019D00024)。Colm O'hUiginとLaboratory of Integrative Cancer Immunology-Microbiome and Genetics CoreのメンバーであるWuxing (Jane) YuanとShah M. Rashedには、16S rRNAライブラリーの前処理、配列決定、一次データの保存に感謝する。Jon Inglefield博士とYanyu Wang博士(Frederick Histopathology Core, Leidos Biomedical Research, Inc.)には、Luminexによる貴重な解析を提供していただいた。Baktiar Karim博士およびTammy Beachley博士、Donna Butcher博士、Tamara Morgan博士を含むFrederick National LaboratoryのMolecular Histopathology Laboratoryのメンバーには、cFOS染色と一次データの保存にご協力いただいた。また、LASP(Laboratory Animal Sciences Program)の動物サポート、およびマウスの繁殖戦略について有益な議論をしてくれたMarina Villamor-Paya博士に感謝する。
データ解析には、NIH HPC Biowulfクラスタ(http://hpc.nih.gov)、NCI Partek Flowワークスペース、NIH Halo Image Analysis Resourceの計算リソースやストレージを利用した。グラフィカルな図はBiorender (http://biorender.com/)で作成した。
参考文献
1.↵ J. M. Kocarnik、K. Compton、F. E. Dean、W. Fu、B. L. Gaw、J. D. Harvey、H. J. Henrikson、D. Lu、A. Pennini、R. Xu、E. Ababneh、M. Abbasi-Kangevari、H. Abbastabar、S. M. Abd-Elsalam、A. Abdoli、A. Abedi、H. Abidi、H. Abolhassani、I. A. アデデジ、Q. E. S. Adnani, S. M. Advani, M. S. Afzal, M. Aghaali, B. O. Ahinkorah, S. Ahmad, T. Ahmad, A. Ahmadi, S. Ahmadi, T. Ahmed Rashid, Y. Ahmed Salih, G. T. Akalu, A. Aklilu, T. Akram, C. J. Akunna, H. Al Hamad, F. Alahdab, Z. アル・アリ、S.アリ、Y.アリモハマディ、V. Alipour、S. M. Aljunid、M. Alkhayyat、A. Almasi-Hashiani、N. A. Almasri、S. A. A. Al-Maweri、S. Almustanyir、N. Alonso、N. Alvis-Guzman、H. Amu、E. W. Anbesu、R. Ancuceanu、F. アンサリ、A.アンサリ-モガダム、M.H.アントウィ、D.アンヴァリ、A.E.アニャソドー、M.アキール、J.アラブルー、M.アラブ-ゾザニ、O.アレム、H.アリフィン、T.アリポフ、M.アルシャド、A.アルタマン、J.アルラッパン、Z. Asemi、M. Asghari Jafarabadi、T. Ashraf、P. Atorkey、A. Aujayeb、M. Ausloos、A. F. Awedew、B. P. Ayala Quintanilla、T. Ayenew、M. A. Azab、S. Azadnajafabad、A. Azari Jafari、G. アザリアン、A.Y.アッザム、A.D.バディエ、S.バハドリー、A.A.バイグ、J.L.ベイカー、S.バラクリシュナン、M.バナッハ、T.W.バーニガウゼン、F.バローン・アデシ、F.バラ、A.バロー、M.ベーザディファル、U.I. ベルガウミ、W.M.M.ベザベ、Y.M.ベザビ、D.S.バガット、A.S.バガヴァトゥーラ、N.バルドワジ、P.バルドワジ、S.バスカール、K.バッタチャリヤ、V. S.Bhojaraja、S.Bibi、A.Bijani、A.Biondi、C.Bisignano、T.Bjørge、A.Bleyer、O.Blyuss、O.A.Bolarinwa、S.R.Bolla、D.Braithwaite、A.Blar、H.Brenner、M.T.Bustamante-Teixeira、N.S.Butt、Z. A.バット、F.L.カエターノ・ドス・サントス、Y. Cao、G. Carreras、F. Catalá-López、F. Cembranel、E. Cerin、A. Cernigliaro、R. C. Chakinala、S. K. Chattu、V. K. Chattu, P. Chaturvedi, O. Chimed-Ochir, D. Y. チョー、D.J.クリストファー、D.-T. チュー、M.T.チョン、J.コンデ、S.コルテス、P.A.コルテシ、V. M. コスタ、A.R.クーニャ、O.ダドラス、A.B.ダグニュー、S.M.A.ダーラウィ、X. Dai、L. Dandona、R. Dandona、A. M. Darwesh、J. das Neves、F. P. De la Hoz、A. B. Demis、E. Denova-Gutiérrez、D. Dhamnetiya、M. L. Dhimal、M. Dhimal、M. Dianatinasab、D. Diaz、S. Djalalinia、H. P. Do、S. Doaei、F. Dorostkar、F. W. dos Santos Figueiredo、T. R. Driscoll、H. Ebrahimi、S. Eftekharzadeh、M. El Tantawi、H. El-Abid、I. エルバラジ、H. R. エルハバシー、M. エルハディ、S. I. エル=ジャファリー、B.エシュラティ、S.エスカンダリエ、F.エスメイルザデ、A.エテマディ、S.エジコウリ、M.ファイサルディン、E.J.A.ファラオン、J.ファレス、F.ファルザドファル、A.H.フェロゼ、S.フェレロ、L.フェロ・デシデリ、I. フィリプ、F.フィッシャー、J.L.フィッシャー、M.フォロタン、福本哲郎、P.A.ガール、M.M.ガッド、M.A.ガダーニャ、S.ガルス、M.ギャスパー・フォンセカ、A.ゲタチュー・オサ、M.ガフーリファード、A.ガシュゲー、N. Ghith、M. Gholamalizadeh、S. A. Gilani、T. G. Ginindza、A. T. T. Gizaw、J. C. Glasbey、M. Golechha、P. Goleij、R. S. Gomez、S. V. Gopalani、G. Gorini、H. Goudarzi、G. Grosso、M. I. M.グバリ、M.R.ゲラ、A.グハ、D.S.グナセケラ、B.グプタ、V. B. グプタ、V.K.グプタ、R.A.グティエレス、N.ハフェジ=ネジャド、M.R.ハイダー、A.ハジ=ミルザイアン、R.ハルワニ、R.R.ハマデ、S.ハメード、S.ハミディ、A.ハニフ、S.ハケ、N.I. ハーリアント、J. M. ハロー、A. I. ハサバラ、S.ハッサニプール、R.J.ヘイ、S.I. ヘイ、K.ハヤト、G.ハイダリ、M.ハイダリ、B.Y.エレラ=セルナ、C.ヘルテリウ、K.ヘザム、R.ホラ、M.M.ホサイン、M.B.H.ホサイン、M.Hosseini, M. Hosseini, M. Hosseinzadeh, M. Hostiuc, S. Hostiuc, M. Househ, M. Hsairi, J. Huang, F. N. Hugo, R. Hussain, N. R. Hussein, B.-F. ファン、I. Iavicoli、S. E. Ibitoye、F. Ida、K. S. Ikuta、O. S. Ilesanmi、I. M.イリッチ、M.D.イリッチ、L.M.イルハム、J.Y.イスラム、R.M.イスラム、S.M.S.イスラム、N.E.イスマイル、G.イソラ、M.岩上、L.ジェイコブ、V. Jain, M. B. Jakovljevic, T. Javaheri, S. Jayaram, S. B. Jazayeri, R. P. Jha, J. B. Jonas, T. Joo, N. Joseph, F. Joukar, M. Jürisson, A. Kabir, D. Kahrizi, L. R. Kalankesh, R. Kalhor, F. Kaliyadan, Y. Kalkonde, A. Kamath, N. Kameran Al-Salihi, H. Kandel, N. Kapoor, A. Karch, A. S. Kasa, S. V. Katikireddi, J. H. Kauppila, T. Kavetskyy, S. A. Kebede, P. Keshavarz, M. Keykhaei, Y. S. Khader, R. Khalilov, G. Khan, M. Khan, M. N. Khan, M. A. B. Khan, Y.-. H. Khang, A. M. Khater, M. Khayamzadeh, G. R. Kim, Y. J. Kim, A. Kisa, S. Kisa, K. Kissimova-Skarbek, J. A. Kopec, R. Koteeswaran, P. A. Koul, S. L. Koulmane Laxminarayana, A. Koyanagi, B. Kucuk Bicer, N. Kugbey, G. A. Kumar, N. Kumar, O. P. Kurmi, T. Kutluk, C. La Vecchia, F. H. Lami, I. ランディレス、P.ラウリオラ、S.リー、S.W.H.リー、W.-C. リー、Y.H.リー、J.リー、E.レオン、J.リー、M.-C. リー、X.Liu、J. A. Loureiro、R. Lunevicius、M. Magdy Abd El Razek、A. Majeed、A. Makki、S. Male、A. A. Malik、M. A. Mansournia、S. Martini、S. Z. Masoumi、P. Mathur、M. McKee、R. Mehrotra、W. Mendoza、R. G. Menezes、E. W. Mengesha、M. K. Mesregah、T. Mestrovic、J. Miao Jonasson、B. Miazgowski、T. Miazgowski、I. M. Michalek, T. R. Miller, H. Mirzaei, H. R. Mirzaei, S. Misra, P. Mithra, M. Moghadaszadeh, K. A. Mohammad, Y. Mohammad, M. Mohammadi, S. M. Mohammadi, A. Mohammadian-Hafshejani, S. Mohammed, N. Moka, A. H. H. Mokdad、M. Molokhia、L. Monasta、M. A. Moni、M. A. Moosavi、Y. Moradi、P. Moraga、J. Morgado-da-Costa、S. D. Morrison、A. Mosapour、S. Mubarik、L. Mwanri、A. J. Nagarajan、S. P. Nagaraju、C. Nagata、M. D. Naimzada、V. Nangia, A. A. Naqvi, S. Narasimha Swamy, R. Ndejjo, S. O. Nduaguba, I. ネゴイ、S.M.ネグル、S.ノイパネ・カンデル、C.T.グエン、H.L.T.グエン、R.K.ニアジ、C.A.ンナジ、N.M.ヌール、V. ヌニェス・サムーディオ,C.Nzoputam, B. Oancea, C. Ochir, O. Odukoya, F. A. Ogbo, A. T. Olagunju, B. O. Olakunde, E. Omar, A. Omar Bali, A. E. E. Omonisi, S. Ong, O. E. Onwujekwe, H. Orru, D. V. Ortega-Altam. V. Ortega-Altamirano、N. Otstavnov、S. S. Otstavnov、M. O. Owolabi、M. P. J. R. Padubidri、K. Pakshir、A. Pana、D. Panagiotakos、S. Panda-Jonas、S. Pardhan、E. C. パク、E.Park、F. Pashazadeh Kan、H. K. Patel、J. R. Patel、S. Pati、S. M. Pattanshetty、U. Paudel、D. M. Pereira、R. B. Pereira、A. Perianayagam、J. D. Pillay、S. Pirouzpanah、F. Pishgar、I. Podder、M. J. Postma、H. Pourjafar、A. Prashant、L. Preotescu、M. Rabiee、N. Rabiee、A. Radfar、R. A. Radhakrishnan、V. ラハクリシュナン、A.ラフィー、F.ラヒム、S.ラヒムザデ、M.ラフマン、M.A.ラフマン、A.M.ラフマニ、N.ラジャイ、A.ラジェッシュ、I. ラコヴァチ、P.ラム、K.ラメザンザデ、K.ラナバット、P.ラナシンヘ、C.R.ラオ、S.J.ラオ、R.ラワシザデ、M.S.ラゼギニア、A.M.N.レンザホ、N. Rezaei、N. Rezaei、A. Rezapour、T. J. Roberts、J. A. B. Rodriguez、P. Rohloff、M. Romoli、L. Ronfani、G. Roshandel、G. M. Rwegerera、M. S. S、 Sabour、B. Saddik、U. Saeed、A. Sahebkar、H. Sahoo、S. Salehi、M. R. Salem、H. Salimzadeh、M. Samaei、A. M. Samy、J. Sanabria、S. Sankararaman、M. M. Santric-Milicevic、Y. Sardiwalla、A. Sarveazad、B. Sathian、M. Sawhney、M. Saylan、I. J.C.シュナイダー、M.セケリヤ、A.セイラニ、O.シャファート、Z. Shaghaghi, M. A. Shaikh, E. Shamsoddin, M. Shannawaz, R. Sharma, A. Sheikh, S. Sheikhbahaei, A. Shetty, J. K. Shetty, P. H. Shetty, K. Shibuya, R. Shirkoohi, K. M. Shivakumar, V. Shivarov, S. Si. Shivarov、S. Siabani、S. K. Siddappa Malleshappa、D. A. S. Silva、J. A. Singh、Y. Sintayehu、V. Y. Skryabin, A. A. Skryabina, M. J. Soeberg, A. Sofi-Mahmudi, H. Sotoudeh, P. Steiropoulos, K. Straif, R. Subedi, M. B. Sufiyan, I. スルタン、S.スルタナ、D.スール、V. Szerencs、M. Szócska、R. Tabarés-Seisdedos、T. Tabuchi、H. Tadbiri、A. Taherkhani、K. Takahashi、I. M.タラート、K.-K. タン、V.Y. Tat、B. A. A. Tedla、Y. G. Tefera、A. Tehrani-Banihashemi、M.-H. Temsah、F. H. Tesfay、G. A. Tessema、R. Thapar、A. Thavamani、V. トグルーヴァ・チャンドラセカール、N. トーマス、H. R. トヒディニク、M. トゥヴィエ、M. R. トヴァニ=パローネ、E. トライニ、B. X. トラン、K. B. トラン、M. T. N. トラン、J. P. トリパシー、B. S. トゥサ、I. ウラ、S. ウラ、K. K. ウマパティ、B. ウニクリシュナン、E. ウパディヤイ、M. ヴァカンテ、M. ヴァエジ、S. ヴァラダン・タフバズ、D. Z. ヴェラスケス、M.ヴェルー、F.S.ヴィオランテ、V. ヴラソフ、B. ヴォ、V. Volovici、G. T. Vu、Y. Waheed、R. G. Wamai、P. Ward、Y. F. Wen、R. Westerman、A. S. Winkler、L. Yadav、S. H. Yahyazadeh Jabbari、L. Yang、S. Yaya、T. S. Y. Yazie、Y. Yeshaw、N. Yonemoto、M. Z. Younis, Z. ユセフィ、C. ユー、D. ユース、I. ユヌサ、V. Zadnik, F. Zare, M. S. Zastrozhin, A. Zastrozhina, J. Zhang, C. Zhong, L. Zhou, C. Zhu, A. Ziapour, I. R.Zimmermann、C.Fitzmaurice、C.J.L.Murray、L.M.Force、2010年から2019年までの29の癌グループの癌罹患率、死亡率、生命喪失年数、障害とともに生きた年数、障害調整生存年。JAMA Oncol. 8, 420-444 (2022).Google Scholar
2.↵ H. Rumgay, M. Arnold, J. Ferlay, O. Lesi, C. J. Cabasag, J. Vignat, M. Laversanne, K. A. McGlynn, I. Soerjomataram、2020年における原発性肝がんの世界的負担と2040年までの予測。J. Hepatol. 77, 1598-1606 (2022).CrossRefGoogle Scholar
3.↵ D. X. Nguyen, P. D. Bos, J. Massagué, Metastasis: from dissemination to organ-specific colonization. Nat. Cancer 9, 274-284 (2009).CrossRefPubMedWeb of ScienceGoogle Scholar
4.↵ K. J. Hiam-Galvez, B. M. Allen, M. H. Spitzer, 癌における全身性免疫。Nat. Cancer 21, 345-359 (2021).CrossRefPubMedGoogle Scholar
5.↵ M. Zheng, Z. Tian, Liver-Mediated Adaptive Immune Tolerance. Front. Immunol. 10 (2019).Google Scholar
6.↵ M. W. Robinson, C. Harmon, C. O'Farrelly, Liver immunology and its role in inflammation and homeostasis. Cell. 免疫学 Immunol. 13, 267-276 (2016).CrossRefPubMedGoogle Scholar
7.↵ R. Bai, N. Chen, L. Li, N. Du, L. Bai, Z. Lv, H. Tian, J. Cui, Mechanisms of Cancer Resistance to Immunotherapy. Front. Oncol. 10, 1290 (2020).PubMedGoogle Scholar
8. B. Ruf、B. Heinrich、T. F. Greten、肝細胞癌の免疫生物学と免疫療法:自然免疫細胞と自然免疫様免疫細胞にスポットライトをあてて。Cell. Cell. Mol. Immunol. 18, 112-127 (2021).PubMedGoogle Scholar
9.ȕ T. F. Greten、A. Villanueva、F. Korangy、B. Ruf、M. Yarchoan、L. Ma、E. Ruppin、X. W.Wang、肝細胞癌の免疫療法のためのバイオマーカー。Nat. Rev. Clin. Oncol., 1-19 (2023).Google Scholar
10.↵ A. H. Zahalka, P. S. Frenette, Nerves in cancer. Nat. Cancer 20, 143-157 (2020).CrossRefPubMedGoogle Scholar
11.↵ H. D. Reavis, H. I. Chen, R. Drapkin, Tumor Innervation: 癌にも神経がある。Trends Cancer 6, 1059-1067 (2020).Google Scholar
12.↵ H. S. Venkatesh, T. B. Johung, V. Caretti, A. Noll, Y. Tang, S. Nagaraja, E. M. Gibson, C. W. Mount, J. Polepalli, S. S. Mitra, P. J. Woo, R. C. Malenka, H. Vogel, M. Bredel, P. Mallick, M. Monje, Neuronal Activity Promotes Glioma Growth through Neuroligin-3 Secretion. Cell 161, 803-816 (2015).CrossRefPubMedGoogle Scholar
13. H. S. Venkatesh, L. T. Tam, P. J. Woo, J. Lennon, S. Nagaraja, S. M. Gillespie, J. Ni, D. Y. Duveau, P. J. Morris, J. J. Zhao, C. J. Thomas, M. Monje, Targeting neuronal activity-regulated neuroligin-3 dependency in high grade glioma. Nature 549, 533-537 (2017).CrossRefPubMedGoogle Scholar
14.ȕ H. S. Venkatesh, W. Morishita, A. C. Geraghty, D. Silverbush, S. M. Gillespie, M. Arzt, L. T. Tam, C. Espenel, A. Ponnuswami, L. Ni, P. J. Woo, K. R. Taylor, A. Agarwal, A. Re. アガルワル、A. レゲヴ、D. ブラング、H. フォーゲル、S. ハーヴェイ=ジャンパー、D. E. バーグルス、M. L. スヴァ、R. C. マレンカ、M. モンジェ、神経膠腫の神経回路への電気的およびシナプス的統合。Nature 573, 539-545 (2019).CrossRefPubMedGoogle Scholar
15.↵ J. R. Huh, H. Veiga-Fernandes, Neuroimmune circuits in inter-organ communication. Nat. Rev. Immunol. 20, 217-228 (2020).CrossRefGoogle Scholar
16.↵ M. Monje、J. C. Borniger、N. J. D'Silva、B. Deneen、P. B. Dirks、F. Fattahi、P. S. Frenette、L. Garzia、D. H. Gutmann、D. Hanahan、S. L. Hervey-Jumper、H. Hondermarck、J. B. Hurov、A. Kepecs、S. M. Knox、A. C. Lloyd、C. Magnn. Lloyd, C. Magnon, J. L. Saloman, R. A. Segal, E. K. Sloan, X. Sun, M. D. Taylor, K. J. Tracey, L. C. Trotman, D. A. Tuveson, T. C. Wang, R. A. White, F. Winkler, Roadmap for the Emerging Field of Cancer Neuroscience. Cell 181, 219-222 (2020).CrossRefGoogle Scholar
17.ȕ K. C. Bauer, K. E. Huus, B. B. Finlay, Microbes and the mind: Emerging hallmarks of the gut microbiota-brain axis. Cell. Microbiol. 18, 632-644 (2016).CrossRefGoogle Scholar
18.ȕ K. N. Browning, S. Verheijden, G. E. Boeckxstaens, The Vagus Nerve in Appetite Regulation, Mood, and Intestinal Inflammation. Gastroenterology 152, 730-744 (2017).CrossRefPubMedGoogle Scholar
19.↵神谷明彦、葉山雄一郎、加藤慎一郎、下村淳一、入江啓一郎、金子理沙、柳川恭子、小林恭子、落谷恒夫、自律神経線維の神経支配と活性の遺伝子操作と乳がん進行への影響。Nat. Neurosci. 22, 1289-1305 (2019).Google Scholar
20.↵ Z. Dubeykovskaya、Y. Si、X. Chen、D. L. Worthley、B. W. Renz、A. M. Urbanska、Y. Hayakawa、T. Xu、C. B. Westphalen、A. Dubeykovskiy、D. Chen、R. A. Friedman、S. アスファハ、K.ナガー、Y.テーラー、S.ムトゥパラニ、J.G.フォックス、J.キタジェフスキー、T.C.ワン、神経支配は脾臓TFF2を刺激し、骨髄細胞の膨張と癌を阻止する。Nat. Commun. 7, 10517 (2016).Google Scholar
21.↵ L. Wang, J. Xu, Y. Xia, K. Yin, Z. Li, B. Li, W. Wang, H. Xu, L. Yang, Z. Xu, Muscarinic acetylcholine receptor 3 mediates vagus nerve-induced gastric cancer. Oncogenesis 7, 1-9 (2018).CrossRefGoogle Scholar
22.↵ B. W. Renz, T. Tanaka, M. Sunagawa, R. Takahashi, Z. Jiang, M. Macchini, Z. Dantes, G. Valenti, R. A. White, M. A. Middelhoff, M. Ilmer, P. E. Oberstein, M. K. Angele, H. Deng, Y. Hayakawa, C. B. Westphalen, J. Werner, H. Remotti、M. Reichert、Y. H. Tailor、K. Nagar、R. A. Friedman、A. C. Iuga、K. P. Olive、T. C. Wang、ムスカリン受容体を介したコリン作動性シグナルは、直接的および間接的に膵臓腫瘍の発生と癌幹細胞化を抑制する。Cancer Discov. 8, 1458-1473 (2018).Abstract/FREE Full TextGoogle Scholar
23.ț C. Magnon, S. J. Hall, J. Lin, X. Xue, L. Gerber, S. J. Freedland, P. S. Frenette, Autonomic Nerve Development Contributes to Prostate Cancer Progression. Science 341, 1236361 (2013).Google Scholar
24.↵ S.-C. Wu, H.-T. チェン、Y.-C. Wang, C.-W. Tzeng, C.-H. Hsu, C.-H. 呉宏貞(Wu, H.-T. Chen), Y.-C. Wang, C.-W. Tzeng, C.-H. Hsu, C.-H. Muo, Decreased risk of liver and intrahepatic cancer in non-H. pylori infection perforated peptic ulcer patients with truncal vagotomy: a nationwide study. Sci. Rep. 11, 15594 (2021).Google Scholar
25.↵ C. Zheng、B. E. Snow、A. J. Elia、R. Nechanitzky、C. Dominguez-Brauer、S. Liu、Y. Tong、M. A. Cox、E. Focaccia、A. C. Wakeham、J. Haight、C. Tobin、K. Hodgson、K. T. Gill、W. Ma, T. Berger, M. Heikenwälder, M. E. Saunders, J. Fortin, S. Y. Leung, T. W. Mak, Tumor-specific cholinergic CD4+ T lymphocytes guide immunosurveillance of hepatocellular carcinoma. Nat. Cancer, 1-18 (2023).Google Scholar
26.↵ L. Zhang, L. L. Wu, H. B. Huan, X. J. Chen, X. D. Wen, D. P. Yang, F. Xia, 肝細胞癌における交感神経と副交感神経の支配。Neoplasma 64, 840-846 (2017).Google Scholar
27.ȕ L. V. Borovikova, S. Ivanova, M. Zhang, H. Yang, G. I. Botchkina, L. R. Watkins, H. Wang, N. Abumrad, J. W. Eaton, K. J. Tracey, Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 405, 458-462 (2000).CrossRefPubMedWeb of ScienceGoogle Scholar
28.↵ V. A. Pavlov, K. J. Tracey, The vagus nerve and the inflammatory reflex-linking immunity and metabolism. Nat. Endocrinol. 8, 743-754 (2012).CrossRefPubMedGoogle Scholar
29.↵ 寺谷哲也, 三上洋一, 中本直樹, 鈴木崇司, 原田祐輔, 岡林恭一郎, 萩原義人, 谷木直樹, 河野和典, 柴田聡, 宮本和彦, 石亀秀樹, P.-S. Chu, T. Sujino, W. Suda, M. Hattori, M. Matsui, T. Okada, H. Okano, M. Inoue, T. Yada, Y. Kitagawa, A. Yoshimura, M. Tanida, M. Tsuda, Y. Iwasaki, T. Kanai, The liver-brain- gut neural arc maintains the Treg cell niche in the gut. Nature 585, 591-596 (2020).CrossRefGoogle Scholar
30.↵ A. Pocai, S. Obici, G. J. Schwartz, L. Rossetti, A brain-liver circuit regulates glucose homeostasis. Cell Metab. 1, 53-61 (2005).CrossRefPubMedWeb of ScienceGoogle Scholar
31.↵ W. E. Naugler, T. Sakurai, S. Kim, S. Maeda, K. Kim, A. M. Elsharkawy, M. Karin, Gender Disparity in Liver Cancer Due to Sex Differences in MyD88-Dependent IL-6 Production. Science 317, 121-124 (2007).Abstract/FREE Full TextGoogle Scholar
32.ț B. Ruf, V. V. Catania, S. Wabitsch, C. Ma, L. P. Diggs, Q. Zhang, B. Heinrich, V. Subramanyam, L. L. Cui, M. Pouzolles, C. N. Evans, R. Chari, S. Sakai, S. Oh, C. E. Barry III、 D.L.バーバー、T.F.グレーテン、粘膜関連不変T細胞の活性化は広範な抗腫瘍反応を誘導する。Cancer Immunol. Res. 9, 1024-1034 (2021).Abstract/FREE Full TextGoogle Scholar
33.↵ C. Ma、M. Han、B. Heinrich、Q. Fu、Q. Zhang、M. Sandhu、D. Agdashian、M. Terabe、J. A. Berzofsky、V. Fako、T. Ritz、T. Longerich、C. M. Theriot、J. A. McCulloch、S. Roy、W. Yuan、V. Thovaroch、S. Roy、W. 袁、V. Thovarai、S. K. Sen、M. Ruchirawat、F. Korangy、X. W. Wang、G. Trinchieri、T. F. Greten、腸内細菌が介在する胆汁酸代謝は、NKT細胞を介して肝臓がんを制御する。Science 360, eaan5931 (2018).Abstract/FREE Full TextGoogle Scholar
34.↵ S. Wabitsch, J. D. McCallen, O. Kamenyeva, B. Ruf, J. C. McVey, J. Kabat, J. S. Walz, Y. Rotman, K. C. Bauer, A. J. Craig, M. Pouzolles, I. Phadke, V. Catania, B. L. Green, C. Fu, L. B. L. Green、C. Fu、L. P. Diggs、B. Heinrich、X. W. Wang、C. Ma、T. F. Greten、メトホルミン投与は、NAFLDマウスにおける免疫チェックポイント阻害剤治療に対するCD8+ T細胞応答を回復させる。J. Hepatol. 77, 748-760 (2022).Google Scholar
35.↵ D. Aran, R. Camarda, J. Odegaard, H. Paik, B. Oskotsky, G. Krings, A. Goga, M. Sirota, A. J. Butte, Comprehensive analysis of normal adjacent to tumor transcriptomes. Nat. Commun. 8, 1077 (2017).CrossRefPubMedGoogle Scholar
36.↵ B. Ruf, M. Bruhns, S. Babaei, N. Kedei, L. Ma, M. Revsine, M.-R. Benmebarek、C. Ma、B. Heinrich、V. Subramanyam、J. Qi、S. Wabitsch、B. L. Green、K. C. Bauer、Y. Myojin、L. T. Greten、J. D. McCallen、P. Huang、R. Trehan、X. Wang、A. Nur、D. Q. M. Soika、M. Pouzolles、C. N. Evans、R. Chari、D. E. Kleiner、W. Telford, K. Dadkhah, A. Ruchinskas, M. K. Stovroff, J. Kang, K. Oza, M. Ruchirawat, A. Kroemer, X. W. Wang, M. Claassen, F. Korangy, T. F. Greten, Tumor-associated macrophage trigger MAIT cell dysfunction at the HCC invasive margin. Cell 186, 3686-3705.e32 (2023).Google Scholar
37.↵ M. J. Kelly, C. Breathnach, K. J. Tracey, S. C. Donnelly, Manipulation of the inflammatory reflex as a therapeutic strategy. Cell Rep. Med. 3, 100696 (2022).Google Scholar
38.ȕ S. Garofalo, G. Cocozza, A. Mormino, G. Bernardini, E. Russo, D. Ielpo, D. Andolina, R. Ventura, K. Martinello, M. Renzi, S. Fucile, M. Laffranchi, E. P. Mortari, R. Carsetti, G. Sciumè, S. Sozzani, A. Santoni, M.-E. ナチュラルキラー細胞と自然リンパ球1は、インターフェロン-γとアセチルコリンを介して、マウスの不安様行動と記憶を制御する。Nat. Commun. 14, 3103 (2023).Google Scholar
39.↵ C. Zheng, L. Zheng, J.-K. 郭赫、張曜、郭旭、姜斌、胡梨央、黄毓、張琪、劉泽、董睦、胡旭、欧陽、彭贺、張泽、単一細胞シークエンシングで明らかになった肝癌浸潤T細胞のランドスケープ。Cell 169, 1342-1356.e16 (2017).CrossRefPubMedGoogle Scholar
40.↵ G. A. Smith, J. Taunton, A. Weiss, IL-2Rβの存在量は、CD4+およびCD8+ T細胞におけるIL-2シグナル動態を異なって調整する。シグナル。10, eaan4931 (2017).Abstract/FREE Full TextGoogle Scholar
41.ȕ Y.-Y. このような研究は、日本免疫学会(J. Immunol. J. Immunol. Baltim. Md 1950 189, 1400-1405 (2012).Google Scholar
42.↵ T. Sekine, A. Perez-Potti, S. Nguyen, J.-B. Gorin, V. H. Wu, E. Gostick, S. Llewellyn-Lacey, Q. Hammer, S. Falck-Jones, S. Vangeti, M. Yu, A. Smed-Sörensen, A. Gaballa, M. Uhlin, J. K. Sandberg, C. Brander、P. Nowak、P. A. Goepfert、D. A. Price、M. R. Betts、M. Buggert、TOXは、疲弊し多機能化したヒトエフェクターメモリーCD8+T細胞に発現している。Sci. Immunol. 5, eaba7918 (2020).Abstract/FREE Full TextGoogle Scholar
43.↵ M. セイファート、M.-R. Benmebarek、D. Briukhovetska、F. Märkl、J. Dörr、B. L. Cadilha、J. Jobst、S. Stock、D. Andreu-Sanz、T. Lorenzini、R. Grünmeier、A. Oner、H. Obeck、L. Majed、D. Dhoqina、M. Feinendegen、A. A2ARとA2BRの選択的二重拮抗薬AB928/etrumadenantがCAR T細胞の機能に与える影響。Br. J. Cancer 127, 2175-2185 (2022).Google Scholar
44.↵ C. Trapnell, D. Cacchiarelli, J. Grimsby, P. Pokharel, S. Li, M. Morse, N. J. Lennon, K. J. Livak, T. S. Mikkelsen, J. L. Rinn, The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 32, 381-386 (2014).CrossRefPubMedGoogle Scholar
45.↵ C.-M. Zhao、Y. Hayakawa、Y. Kodama、S. Muthupalani、C. B. Westphalen、G. T. Andersen、A. Flatberg、H. Johannessen、R. A. Friedman、B. W. Renz、A. K. Sandvik、V. Beisvag, H. Tomita, A. Hara, M. Quante, Z. Li, M. D. Gershon, K. Kaneko, J. G. Fox, T. C. Wang, D. Chen, Denervation suppresses gastric tumorigenesis. Sci. Transl. Med. 6, 250ra115 (2014).Abstract/FREE Full TextGoogle Scholar
46.↵M. E. Schmidt、S. Hermann、V. Arndt、K. Steindorf、15種類のがんにおける長期的な身体的・感情的・認知的疲労の有病率と重症度。Cancer Med. 9, 8053-8061 (2020).Google Scholar
47. J. M. Llovet, S. Ricci, V. Mazzaferro, P. Hilgard, E. Gane, J.-F. Blanc, A. C. de Oliveira, A. Santoro, J.-L. Raoul, A. Forner, M. Schwartz, C. Porta, S. Zeuzem, L. Bolondi, T. F. Greten, P. R. Galle, J.-F. 進行肝細胞癌におけるソラフェニブ。N. Engl. 359, 378-390 (2008).CrossRefPubMedWeb of ScienceGoogle Scholar
48.ȕ B. Wolff, S. A. Raheem, S. A. Alshawi, J. M. Regan, L. R. Feng, L. N. Saligan, 雄マウスの骨盤照射による疲労様行動の誘発は、認知行動とBDNF発現を変化させる。PLOS ONE 15, e0235566 (2020).Google Scholar
49.↵ S. Fan, C. Eiser, M. Ho, Hepatocellular Carcinoma Patients in Health-Related Quality of Life: A Systematic Review. Clin. Gastroenterol. Hepatol. 8, 559-564.e10 (2010).CrossRefPubMedGoogle Scholar
50.↵ N. R. Gessner, M. Peiravi, F. Zhang, S. Yimam, D. Springer, S. T. Harbison, A conserved role for frizzled in sleep architecture. SLEEP Adv. 4, zpad045 (2023).Google Scholar
51.ȕ K. C. Bauer, E. M. York, M. S. Cirstea, N. Radisavljevic, C. Petersen, K. E. Huus, E. M. Brown, T. Bozorgmehr, R. Berdún, L.-. P.ベルニエ, A.H.Y.リー, S.E.ウッドワード, Z.クレクノ, J.ハン, R.E.W.ハンコック, V.アヤラ, B.A.マクビカー, B.B.フィンレイ, 腸内微生物が栄養不良時のミクログリアと認知機能を形成する。Glia 70, 820- 841 (2022).Google Scholar
52.↵ L. Pedersen、M. Idorn、G. H. Olofsson、B. Lauenborg、I. Nookaew、R. H. Hansen、H. H. Johannesen、J. C. Becker、K. S. Pedersen、C. Dethlefsen、J. Nielsen、J. Gehl、B. K. Pedersen, P. thor Straten, P. Hojman, Voluntary Running Suppresses Tumor Growth through Epinephrine and IL-6-Dependent NK Cell Mobilization and Redistribution. Cell Metab. 23, 554-562 (2016).CrossRefPubMedGoogle Scholar
53.↵ B. Bonaz, T. Bazin, S. Pellissier, The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 12, 49 (2018).Google Scholar
54.↵ Y. Minnebo, K. Delbaere, V. Goethals, J. Raes, T. Van de Wiele, K. De Paepe, Gut microbiota response to in vitro transit time variation is mediated by microbial growth rates, nutrient use efficiency and adaptation to in vivo transit time. Microbiome 11, 240 (2023).Google Scholar
55.↵ A.R.ウィルキンソン、D.ジョンストン、人間における食物バリウム食の胃排出と腸通過に及ぼす切断的、選択的、高選択的迷走神経切断の影響。Ann. Surg. 178, 190-193 (1973).CrossRefPubMedGoogle Scholar
56.↵ S. J. Robertson、P. Lemire、H. Maughan、A. Goethel、W. Turpin、L. Bedrani、D. S. Guttman、K. Croitoru、S. E. Girardin、D. J. Philpott、マウスモデルにおける微生物相標準化のための共同飼育法とリターメイト法の比較。Cell Rep. 27, 1910-1919.e2 (2019).CrossRefPubMedGoogle Scholar
57.↵ C. Ma, J. McCallen, J. C. McVey, R. Trehan, K. Bauer, Q. Zhang, B. Ruf, S. Wang, C. W. Lai, G. Trinchieri, J. A. Berzofsky, F. Korangy, T. F. Greten, CSF-1R+ Macrophage Control the Gut Microbiome-Enhanced Liver Invariant NKT Function through IL-18. J. Immunol. 211, 1099-1107 (2023).Google Scholar
58.↵ S. Ni, Z.-Y. Yao, X. Wei, X. Heng, S.-Y. Qu、X. Zhao、Y.-Y. Qi, P.-Y. 葛、C.-P. Xu、N.-Y. Yang、Y. Cao、H.-X. Zhu, R. Guo, Q.-C. 硫化水素による迷走神経刺激とベルベリンによるミクログリア制御機構を明らかにした。Phytother. Res. 36, 2964-2981 (2022).Google Scholar
59.↵ X. Zhang, O. O. Coker, E. S. Chu, K. Fu, H. C. H. Lau, Y.-X. このような背景のもとで、日本では、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」、「食欲の秋」の4つの季節が到来した。Gut 70, 761-774 (2021).Abstract/FREE Full TextGoogle Scholar
60.↵ D. Davar, A. K. Dzutsev, J. A. McCulloch, R. R. Rodrigues, J.-M. ショーバン、R.M.モリソン、R.N.デブラシオ、C.メンナ、Q.ディン、O.パリアーノ、B.ジディ、S.チャン、J.H.バジャー、M.ヴェチズー、A.M.コール、M.R.フェルナンデス、S.プレスコット、R.G.F.コスタ、A.K.バラジ、A.モルグン、I.ヴイコヴィッチ-Cvijin、H.ワン、A. A.ボルハニ、M.B.シュワルツ、H.M.ダブナー、S.J.エルンスト、A.ローズ、Y.G.ナジャール、Y.ベルカイド、J.M.カークウッド、G.トリンキエリ、H.M.ザロア、糞便微生物移植はメラノーマ患者の抗PD-1療法に対する抵抗性を克服する。Science 371, 595-602 (2021).Abstract/FREE Full TextGoogle Scholar
61.↵ C. N. Spencer、J. L. McQuade、V. Gopalakrishnan、J. A. McCulloch、M. Vetizou、A. P. Cogdill、A. W. Khan、X. Zhang、M. G. White、C. B. Peterson、M. C. Wong、G. Morad、T. Rodgers、J. H. Badger, B. A. Helmink, M. C. Andrews, R. R. Rodrigues, A. Morgun, Y. S. Kim, J. Roszik, K. L. Hoffman, J. Zheng, Y. Zhou, Y. B. Medik, L. M. Kahn, S. Johnson, C. W. Hudgens, K. Wani, P.-. O. ゴードロー、A.L.ハリス、M.A.ジャマル、E.N.バルク、E.ペレス-ギジャロ、C.-P. デイ、G.メルリーノ、B.パズドラック、B.S.ロッホマン、R.A.シュツェパニアク=スローン、R.アローラ、J.アンダーソン、C.M.ゾブニウ、E.ポサダ、E.シルマンズ、J.サイモン、L.E.ヘイデュ、E.M.バートン、L.ワン、M.ダン、K.クライス=ドワイヤー、S.シュナイダー、T.チャップマン、N.A. S. Anang, S. Duncan, J. Toker, J. C. Malke, I. C. Glitza, R. N. Amaria, H. A. Tawbi, A. Diab, M. K. Wong, S. P. Patel, S. E. Woodman, M. A. デイビス、M.I.ロス、J.E.ガーシェンバルト、J.E.リー、P.フー、V.ジェンセン、Y.サミュエルズ、R.ストラウスマン、N.J.アジャミ、K.C.ネルソン、L.ネジ、J.F.ペトロシーノ、P. A.Futreal、A.J.Lazar、J.Hu、R.R.Jenq、M.T.Tetzlaff、Y.Yan、W.S.Garrett、C.Huttenhower、P.Sharma、S.S.Watowich、J.P.Allison、L.Cohen、G.Trinchieri、C.R.Daniel、J.A.Wargo、食物繊維とプロバイオティクスは腸内マイクロバイオームとメラノーマ免疫療法反応に影響を及ぼす。Science 374, 1632-1640 (2021).CrossRefPubMedGoogle Scholar
62.↵ E. M. Park, M. Chelvanambi, N. Bhutiani, G. Kroemer, L. Zitvogel, J. A. Wargo, Targeting the gut and tumor microbiota in cancer. Nat. Med. 28, 690-703 (2022).Google Scholar
63.ȕ K. C. Bauer, T. F. Greten, A gut response: 化学療法の効果を微生物代謝産物で調節する。Immunity 56, 750-752 (2023).Google Scholar
64.↵ 米倉聡、S. テリッセ、C. アルヴェス・コスタ・シルヴァ、A. ラファルジュ、V. イエバ、G. フェレール、A.-G. グベ、J.ファルナー、ラフマール、上田欣也、マンソーリ、ピッツァート、リー、マゼンガ、テレマケ、フィデル、ジャウリン、カルトリー、デロジェ、アグラーベ、ドロイン、オポロン、プジェ、マン、ノウンリスト、A. Neunlist, A. Bessard, L. Aymeric, T. Matysiak-Budnik, J. Bosq, P. Hofman, C. P. M. Duong, S. Ugolini, V. Quiniou, S. Berrard, B. Ryffel, O. Kepp, G. Kroemer, B. Routy, L. Lordello, M.-. A. Bani、N. Segata、F. Y. Yengej、H. Clevers、J.-Y. 癌はβ-アドレナリン受容体に依存したストレスイレオパシーを誘導し、発癌に寄与するディスバイオシスを促進する。Cancer Discov. 12, 1128- 1151 (2022).Google Scholar
65.↵ B. Sangro, P. Sarobe, S. Hervás-Stubbs, I. Melero, Advances in immunotherapy for hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 18, 525-543 (2021).CrossRefPubMedGoogle Scholar
66.↵ S. R. Sharpton, B. Maraj, E. Harding-Theobald, E. Vittinghoff, N. A. Terrault, 非アルコール性脂肪肝疾患における腸内細菌を標的とした治療法:系統的レビュー、メタ解析、メタ回帰。Am. J. Clin. Nutr. 110, 139-149 (2019).Google Scholar
67. 木村和彦, 谷田正樹, 永田直樹, 稲葉陽子, 渡辺裕之, 長島田正樹, 太田哲也, 浅原聡子, 木戸義人, 松本正樹, 利内賢一, 中里正樹, 柴本哲也, S. 金子, 春日正樹, 井上博之, 中心性インスリン作用はニコチン性α7アセチルコリン受容体を介した肝Vagal活性化を抑制することによりクッパー細胞を活性化する. Cell Rep. 14, 2362-2374 (2016).Google Scholar
68. Y. Lei, L. Tang, Q. Chen, L. Wu, W. He, D. Tu, S. Wang, Y. Chen, S. Liu, Z. Xie, H. Wei, S. Yang, B. Tang, Disulfiram ameliorates nonalcoholic steatohepatitis by modulating the gut microbiota and bile acid metabolism. Nat. Commun. 13, 6862 (2022).Google Scholar
69.ȕ K. C. Bauer, P. T. Littlejohn, V. Ayala, A. Creus-Cuadros, B. B. Finlay, Nonalcoholic Fatty Liver Disease and the Gut-Liver Axis: Exploring an Undernutrition Perspective. Gastroenterology 162, 1858-1875.e2 (2022).Google Scholar
70.↵ G. D. Sepich-Poore, L. Zitvogel, R. Straussman, J. Hasty, J. A. Wargo, R. Knight, The microbiome and human cancer. Science 371, eabc4552 (2021).Abstract/FREE Full TextGoogle Scholar
71.↵ E. Pasolli, D. T. Truong, F. Malik, L. Waldron, N. Segata, Machine Learning Meta-analysis of Large Metagenomic Datasets: ツールおよび生物学的洞察。PLOS Comput. Biol. 12, e1004977 (2016).CrossRefGoogle Scholar
72.↵ R. F. Schwabe, T. F. Greten, Gut microbiome in HCC - Mechanisms, diagnosis and therapy. J. Hepatol. 72, 230-238 (2020).CrossRefPubMedGoogle Scholar
73.ȕ A. Sivan, L. Corrales, N. Hubert, J. B. Williams, K. Aquino-Michaels, Z. M. Earley, F. W. Benyamin, Y. Man Lei, B. Jabri, M.-L. Alegre, E. B. Chang, T. F. Gajewski, 通常性ビフィズス菌は抗腫瘍免疫を促進し、抗PD-L1効果を促進する。Science 350, 1084-1089 (2015).Abstract/FREE Full TextGoogle Scholar
74.↵ V. Gopalakrishnan、C. N. Spencer、L. Nezi、A. Reuben、M. C. Andrews、T. V. Karpinets、P. A. Prieto、D. Vicente、K. ホフマン、S.C.ウェイ、A.P.コグディル、L.ザオ、C.W.ハジェンズ、D.S.ハッチンソン、T.マンゾ、M.P.デ・マセド、T.コテチーニ、T.クマール、W.S. Chen、S. M. Reddy、R. S. Sloane、J. Galloway-Pena、H. Jiang、P. L. Chen、E. J. Shpall、K. Rezvani、A. M. Alousi、R. F. Chemaly、S. シェルバーン、L. M. ヴェンス、P. C. オクハイセン、V. B. ジェンセン、A. G. スウェンズ、F. マカリスター、E. M. R. サンチェス、Y. チャン、E. ル・シャトリエ、L. Zitvogel、N. Pons、J. L. Austin-Breneman、L. E. Haydu、E. M. Burton、J. M. Gardner、E. Sirmans、J. Hu、A. J. Lazar、T. Tsujikawa、A. Diab、H. Tawbi、I. C. Glitza、W. J. Hwu、S. P. Patel、S. E. Woodman、R. N. Amaria、M. A. Davies、J. E. Gershenwald、P. Hwu、J. E. Lee, J. Zhang, L. M. Coussens, Z. A. Cooper, P. A. Futreal, C. R. Daniel, N. J. Ajami, J. F. Petrosino, M. T. Tetzlaff, P. Sharma, J. P. Allison, R. R. Jenq, J. A. Wargo, Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 359, 97-103 (2018).Abstract/FREE Full TextGoogle Scholar
75.↵ K. Liu, L. Yang, G. Wang, J. Liu, X. Zhao, Y. Wang, J. Li, J. Yang, Metabolic stress drives sympathetic neuropathy within the liver. Cell Metab. 33, 666-675.e4 (2021).Google Scholar
76.↵ K. J. Jensen, G. Alpini, S. Glaser, Hepatic Nervous System and Neurobiology of the Liver. Compr. Physiol. 3, 655-665 (2013).Google Scholar
77.↵ C.-X. エネルギー代謝の制御における自律神経肝神経支配の役割。Biochim. Biophys. BBA - Mol. Basis Dis. 1802, 416-431 (2010).Google Scholar
78.↵ A. Khalyfa, S. Punatar, A. Yarbrough, Hepatocellular Carcinoma: Understanding the Inflammatory Implications of the Microbiome. Int. J. Mol. Sci. 23, 8164 (2022).Google Scholar
79.ȕ J. Dahmus, D. L. Kotler, D. M. Kastenberg, C. A. Kistler, The gut microbiome and colorectal cancer: a review of bacterial pathogenesis. J. Gastrointest. Oncol. 9, 769-777 (2018).CrossRefGoogle Scholar
80.↵ C. Fülling, T. G. Dinan, J. F. Cryan, Gut Microbe to Brain Signaling: 迷走神経で何が起こっているのか?Neuron 101, 998-1002 (2019).CrossRefGoogle Scholar
81.ȕ Z. J. Brown, B. Heinrich, T. F. Greten, 非アルコール性脂肪性肝疾患を有するマウスにおける外科的肝内腫瘍注入による同所性肝腫瘍の樹立。Methods Protoc. 1, 21 (2018).Google Scholar
82.↵ L. Wrzosek, D. Ciocan, P. Borentain, M. Spatz, V. Puchois, C. Hugot, G. Ferrere, C. Mayeur, G. Perlemuter, A.-M. ヒトの微生物叢を従来のマウスに移植すると、腸内細菌叢が持続的に再構築される。Sci. Rep. 8, 6854 (2018).CrossRefPubMedGoogle Scholar
83.ȕ E. Bolyen、J. R. Rideout、M. R. Dillon、N. A. Bokulich、C. C. Abnet、G. A. Al-Ghalith、H. Alexander、E. J. Alm、M. Arumugam、F. Asnicar、Y. Bai、J. E. Bisanz、K. Bittinger、A. ブレインロッド、C.J.ブリスローン、C.T.ブラウン、B.J.キャラハン、A.M.カラバロ・ロドリゲス、J.チェイス、E.K.コープ、R.ダ・シルバ、C.ディーナー、P.C.ドーレスタイン、G.M.ダグラス、D.M.デュラル、C.デュヴァレ、C.F. エドワードソン、M.エルンスト、M.エスタキ、J.フーキエ、J.M.ゴーグリッツ、S.M.ギボンズ、D.L.ギブソン、A.ゴンザレス、K.ゴーリック、J.グオ、B.ヒルマン、S.ホームズ、H.ホルステ、C.ハッテンハワー、G.A.ハットリー、S. ヤンセン、A.K.ジャームッシュ、L.ジャン、B.D.ケーラー、K.B.カン、C.R.キーフ、P.ケイム、S.T.ケリー、D.ナイツ、I.コスター、T.コシオレック、J.クレップス、M.G.I.ランギル、J.リー、R.レイ、Y.-。 X. リュー、E.ロフトフィールド、C.ロズポーン、M.マハー、C.マロッツ、B.D.マーティン、D.マクドナルド、L.J.マキバー、A.V.メルニク、J.L.メトカーフ、S.C.モーガン、J.T.モートン、A.T.ナイミー、J.A.ナバス=モリーナ、L.F.ノーシャス、S.B.オルチャニアン、T. Pearson、S. L. Peoples、D. Petras、M. L. Preuss、E. Pruesse、L. B. Rasmussen、A. Rivers、M. S. Robeson、P. Rosenthal、N. Segata、M. Shaffer、A. Shiffer、R. Sinha、S. J. Song、J. R. Spear、A. D. Swafford、L. R.トンプソン、P.J.トーレス、P.トリン、A.トリパティ、P.J.ターンボー、S.ウル=ハサン、J.J.ファンデルフフト、F.バルガス、Y.バスケス=バエサ、E.フォクトマン、M.フォン・ヒッペル、W.ウォルターズ、Y.ワン、M.ワン、J.ウォーレン、K.C. このような背景の下で、研究者らは、微生物学的な知見に基づき、微生物学的なデータサイエンスの可能性を探ることを目的とした研究を行っている。Biotechnol. 37, 852-857 (2019).CrossRefPubMedGoogle Scholar
84.↵ N. Segata, J. Izard, L. Waldron, D. Gevers, L. Miropolsky, W. S. Garrett, C. Huttenhower, Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60 (2011).CrossRefPubMedGoogle Scholar
85.↵山田真人、宮川哲也、A.Duttaroy、山中章、森口哲也、蒔田玲、小川真理、C.J.Chou、B.Xia、J.N.Crawley、C.C.Felder、C.X.Deng、J.Wess、M3ムスカリン性アセチルコリン受容体を欠損したマウスは低食欲性で痩せ型である。Nature 410, 207-212 (2001).CrossRefPubMedWeb of ScienceGoogle Scholar
86.ȕ T. Kapanadze, J. Gamrekelashvili, C. Ma, C. Chan, F. Zhao, S. Hewitt, L. Zender, V. Kapoor, D. W. Felsher, M. P. Manns, F. Korangy, T. F. Greten, Regulation of accumulation and function of myeloid derived suppressor cells in different mouse models of hepatocellular carcinoma. J. Hepatol. 59, 1007-1013 (2013).CrossRefPubMedWeb of ScienceGoogle Scholar
87.↵ S. Kreiter, M. Vormehr, N. van de Roemer, M. Diken, M. Löwer, J. Diekmann, S. Boegel, B. Schrörs, F. Vascotto, J. C. Castle, A. D. Tadmor, S. P. Schoenberger, C. Huber, Ö. Türeci, U. Sahin, 変異型MHCクラスIIエピトープが癌に対する治療的免疫応答を促進する。Nature 520, 692-696 (2015).CrossRefPubMedGoogle Scholar
88.↵ T. Li, J. Fu, Z. Zeng, D. Cohen, J. Li, Q. Chen, B. Li, X. S. Liu, TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 48, W509-W514 (2020).CrossRefPubMedGoogle Scholar
89.↵ P. J. Thul, C. Lindskog, The human protein atlas: ヒトプロテオームの空間マップ。タンパク質科学出版。27, 233-244 (2018).Google Scholar
90.↵ M. Gillespie, B. Jassal, R. Stephan, M. Milacic, K. Rothfels, A. Senff-Ribeiro, J. Griss, C. Sevilla, L. Matthews, C. Gong, C. Deng, T. Varusai, E. Ragueneau, Y. Haider, B. May, V. Shamovsky, J. Weiser, T. Brunson, N. Sanati、L. Beckman、X. Shao、A. Fabregat、K. Sidiropoulos、J. Murillo、G. Viteri、J. Cook、S. Shorser、G. Bader、E. Demir、C. Sander、R. Haw、G. Wu、L. Stein、H. Hermjakob、P. D'Eustachio, The reactome pathway knowledgebase 2022. Nucleic Acids Res. 50, D687-D692 (2022).CrossRefPubMedGoogle Scholar
91.ț B. Heinrich, Z. J. Brown, L. P. Diggs, M. Vormehr, C. Ma, V. Subramanyam, U. Rosato, B. Ruf, J. S. Walz, J. C. McVey, S. Wabitsch, Q. Fu, S. J. Yu, Q. Zhang, C. W. Lai, U. Sahin, T. F. Greten, Steatohepatitis Impairs T-cell-Directed Immunotherapy Against Liver Tumors in Mice. Gastroenterology 160, 331-345.e6 (2021).PubMedGoogle Scholar
92.↵ S. Karlsson, F. Sundler, B. Ahrén, マウスにおけるガストリン放出ペプチドによるインスリン分泌:神経節作用と直接膵島作用。Am. J. Physiol.-Endocrinol. Metab. 274, E124-E129 (1998).PubMedWeb of ScienceGoogle Scholar
93. E. M. Brown, M. Wlodarska, B. P. Willing, P. Vonaesch, J. Han, L. A. Reynolds, M.-C. このような研究は、日本ではまだほとんど行われていない。Nat. Commun. 6, 7806 (2015).CrossRefPubMedGoogle Scholar
トップへ戻る
前へ 次へ
2024年1月25日掲載
PDFダウンロード
印刷/保存オプション
補足資料
Eメール
共有する
引用ツール
COVID-19 SARS-CoV-2のプレプリント(medRxivおよびbioRxivから)
主題領域
癌生物学
主題領域
すべての記事
動物の行動と認知
生化学
生物工学
生物情報学
生物物理学
癌生物学
細胞生物学
臨床試験
発生生物学
生態学
疫学
進化生物学
遺伝学
ゲノミクス
免疫学
微生物学
分子生物学
神経科学
古生物学
病理学
薬理学と毒性学
生理学
植物生物学
科学コミュニケーションと教育
合成生物学
システム生物学
動物学
bioRxiv の臨床研究パイロットプロジェクトが終了し、健康科学専用サーバー medRxiv (submit.medrxiv.org)が開設されたことに伴い、Clinical Trials と Epidemiology のサブジェクトカテゴリーは新規投稿を締め切りました。臨床試験の結果を報告する新規論文は、medRxivへの投稿が必須となりました。ほとんどの疫学論文もmedRxivに投稿されるべきですが、もし論文に健康に関する情報が含まれていない場合、著者は他のbioRxivの主題カテゴリー(例えば、遺伝学や微生物学)に投稿することもできます。
文脈と評価 x
0 0 0 0 0 0 19
この記事が気に入ったらサポートをしてみませんか?