グリホサート(ラウンドアップTM)が腸内細菌叢の組成と機能性に及ぼす影響

メインコンテンツへスキップ
テイラー・アンド・フランシス・オンライン・ホームページ
ログイン|会員登録
カート
腸内微生物
第15巻 2023年-第2号
オープンアクセス
261
閲覧数
0
これまでのCrossRef引用
3
アルトメトリック
聞く
レビュー
グリホサート(ラウンドアップTM)が腸内細菌叢の組成と機能性に及ぼす影響
https://www.tandfonline.com/doi/full/10.1080/19490976.2023.2263935

ローレン・ウォルシュ,コリン・ヒル &R. ポール・ロスORCIDアイコン
論文 2263935|2023年3月14日受理、2023年9月24日受理、オンライン版公開:2023年10月06日
引用
https://doi.org/10.1080/19490976.2023.2263935
クロスマークのロゴクロスマーク
この記事で
記事全文
数字とデータ
参考文献
引用文献
指標
ライセンス
転載と許可
PDF EPUB
XFacebookLinkedInEmail共有
要旨
広域除草剤ラウンドアップTMの有効成分であるグリホサートは、グリホサートの人体への影響に関する矛盾した報告により、数十年にわたり議論されてきた。グリホサートは、植物において芳香族アミノ酸を生成するシキミ経路の5-エノールピルビルシキミ酸-3-リン酸合成酵素(EPSPS)を阻害するが、この経路は哺乳類には存在しないため、除草剤がヒトに影響を及ぼすことはないと考えられている。しかし、多くの研究が、グリホサートへの暴露が人体における様々な障害の発現に関与していることを示唆している。本総説では、特にグリホサート暴露が腸内細菌叢の構成と機能に及ぼす潜在的影響について概説する。個々の腸内細菌叢の構成が健康に大きく影響するという仮説の裏には、エビデンスが構築されつつある。このため、グリホサートが腸内の有益な微生物の増殖を阻害したり、その機能を変化させたりする可能性は、さらなる検討が必要な重要なテーマである。
キーワード:グリホサート腸内細菌叢抗微生物ヒトの健康ラウンドアップTM
背景
グリホサートは、1974年に初めて製造された広域スペクトル除草剤ラウンドアップTMの有効成分である。引用1 2010年、グリホサートは除草剤としての数十年にわたる使用に続いて抗生物質としての特許を取得し、この化合物の優れた抗菌活性が注目された(米国特許第7771736 B2号)。 引用2 グリホサートは米国で最も使用されている除草剤のひとつで、年間2億8,000万ポンドが使用されている。引用3 このように大量に生産された結果、流出やその後の水循環への取り込みを通じて、環境中にグリホサートが広く存在することになった。グリホサートの残留物は、農作物(環境的に獲得され、処理されたもの)引用4、飲料水、引用5、雨、引用6、家畜飼料引用7、大気中などで見つかっている。2015年、グリホサートは世界保健機関(WHO)によって「ヒトに対しておそらく発がん性がある」と再分類された。 Citation10 にもかかわらず、グリホサートは欧州食品安全機関(EFSA)と米国環境保護庁(EPA)により依然として承認されている。
環境中にグリホサートが広く存在することから、残留物の暴露/摂取はヒトにとってリスク要因であり、ヒト腸内細菌叢に影響を及ぼす可能性がある。腸内細菌叢にはさまざまな微生物が生息しており、免疫系の発達や恒常性の維持、引用14 エネルギー代謝の調節、引用15 病原菌に対するコロニー形成抵抗性、引用16 生理活性代謝産物の生産など、宿主に不可欠ないくつかの機能に関連している17。また、皮膚、引用18 肺、引用19、脳など、身体の他の器官の機能とも関連している。
健康な腸内細菌叢の正確な組成を完全に解明することは、マイクロバイオーム間の個体差や種間の高い機能的冗長性のために困難であるが、個人の腸内細菌叢が乱れると病気になる可能性があることが示されている。 引用21 このような現象は「ディスバイオーシス」と呼ばれることもあり、腸の「正常な」コロニー形成に対する変化が疾患と関連している。 Citation25 炎症性腸疾患(IBD)はその一例であり、患者の糞便サンプルにおいて、プロテオバクテリア(Proteobacteria)とバクテロイーダ(Bacteroidota)の増加とバチロタ(Bacillota)の減少が観察されている。 Citation26 同様に、関節リウマチの発症には、プレボテラ属の増加とバクテロイデス属の減少が関連している。Citation26 多発性硬化症患者のサンプルでは、バクテロイデス、フェカリバクテリウム、プレボテラ、ブチリシモナス、コリンセラが増加し、ビフィドバクテリウム、ストレプトコッカス、メタノブレビバクター、アッカーマンシア・ムチニフィラが濃縮されていることが確認されているCitation27。
グリホサートは、トリプトファン、チロシン、フェニルアラニンなどの芳香族アミノ酸を生産するシキミ酸経路の5-エノールピルビルシキミ酸-3-リン酸合成酵素(EPSPS)を阻害する。この経路は植物、菌類、細菌、原生動物、古細菌に存在し、グリホサートを効果的な抗菌剤にしている。ヒトの体内ではトリプトファン、チロシン、フェニルアラニンは生産されないため、これらは食事や腸内微生物による生産を通じて摂取しなければならない。 引用29 ある研究では、100 µMの純粋なグリホサートで処理すると、サトウキビ作物においてトリプトファンが13%、チロシンが59%、フェニルアラニンが77%減少することが示された。
この総説では、グリホサートが人体に及ぼす可能性のある影響について、特に腸内細菌叢と人体への潜在的影響に焦点を当てて議論する。
グリホサートに対する微生物抵抗性のメカニズム
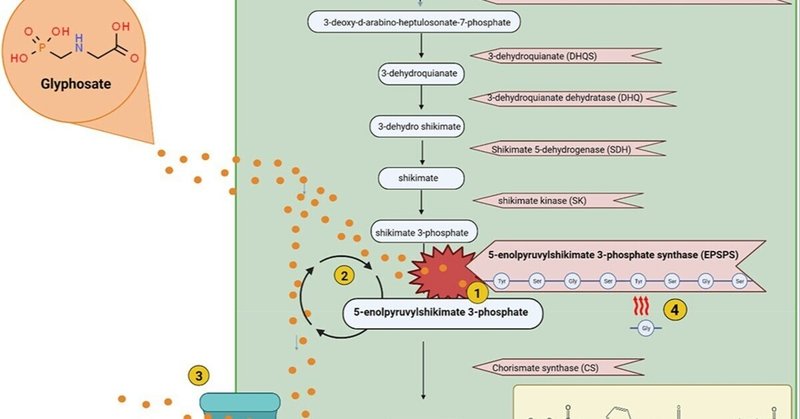
図1に示すように、グリホサートの作用機序は、シキミ酸経路のEPSPSを阻害することであり4、グリホサートがEPSPS酵素に結合することで達成される。その結果、芳香族アミノ酸であるトリプトファン、チロシン、フェニルアラニンの生産が阻害される。
図1. シキミ酸経路とグリホサートに対する抵抗性メカニズム。1. グリホサートはEPSPS酵素を阻害する。2. EPSPS阻害に対抗するためにepsp遺伝子を過剰発現させることができる。3. グリホサートを細胞外に排出するためにエフラックスポンプが使われる。4. EPSPS酵素上のアミノ酸バイオマーカーがグリホサートに対するその酵素の感受性を決定する。アミノ酸配列の変化はグリホサートに対する耐性を与える。Biorenderで作成。
図1. シキメイト経路とグリホサートに対する抵抗性メカニズム。1. グリホサートはEPSPS酵素を阻害する。2. EPSPS阻害に対抗するためにepsp遺伝子を過剰発現させることができる。3. グリホサートを細胞外に排出するためにエフラックスポンプが使われる。4. EPSPS酵素上のアミノ酸バイオマーカーがグリホサートに対するその酵素の感受性を決定する。アミノ酸配列の変化はグリホサートに対する耐性を与える。Biorenderで作成。
フルサイズ表示
細菌の中には、グリホサートの作用を克服するメカニズムを開発した種がある(図1に記載)。そのような耐性メカニズムの一つは標的部位耐性であり、EPSPS活性部位に見られるアミノ酸残基が変化して耐性を付与する。引用33 一部の種が利用するもう一つの耐性メカニズムは非標的部位耐性であり、EPSPS遺伝子の過剰発現が起こるか、グリホサートを細胞外に汲み出すために排出ポンプが利用される。クラスI、II、IVは、EPSPS活性部位における特定のアミノ酸残基の有無によって特徴づけられるという点でよく似ている。これとは対照的に、クラスIIIの酵素にはモチーフが多数存在する。
一般に、放線菌やシュードモナドータに比べ、バチロータはグリホサートに対して耐性がある。大腸菌、サルモネラ腸炎菌、サルモネラ腸炎菌血清型タイフィムリウムのような病原性細菌は、宿主に付随する常在菌に比べて高い耐性を示した。 引用9 グリホサート(グリホサートの40%モノイソプロピルアミン塩溶液とラウンドアップTM LBプラス)の病原性細菌と常在細菌に対する最小発育阻止濃度(MIC)を調べたある研究では、病原性菌株のMIC(20~80 mg/mL)が常在細菌(5~10 mg/mL)よりも有意に高いことがわかった。実際、101の細菌種についてグリホサート感受性/耐性を調べたある研究では、腸内で一般的に見られる細菌種の54%がグリホサートに感受性であった。一方、評価した細菌種の29%は耐性の可能性があった。このような菌種には、Clostridium属、Ruminococcus属、Dorea属のメンバーが含まれる。これらの微生物の増加は、IBDなどの疾患と関連している。引用40 残りの17%の細菌は、分類されていない細菌(10%)と感受性の異なる細菌(7%)で構成されていた。別の研究では、腸内マイクロバイオームに生息する101種890株の細菌を調べた。彼らは異なるタイプのEPSPSをコードする細菌のDNA配列を解析し、腸内細菌叢に存在する細菌の12~26%がグリホサートに対して感受性の可能性があると予測したCitation33。この結果は、中核的な腸内細菌叢におけるグリホサート感受性を確立するために試験された菌株に基づく保守的な計算であり、おそらく2つの研究間の結果の有意な違いを説明している。細菌の中には、グリホサート分解酵素を産生し、グリホサート分解産物(リン酸塩など)を栄養分として利用するように進化したものがある。
2020年に実施された研究では、計算モデリングを用いて細菌に存在するシキミ経路の機能的能力を調べた。引用42 IBDマルチオミクスデータベースの糞便サンプルをインシリコでスクリーニングし、ゲノムレベルと転写レベルの両方でシキミ経路の存在を示唆した。シキミ経路は734のメタゲノムサンプルの98%で検出されたが、対応するメタトランスクリプトームサンプルを解析したところ、シキミ経路を発現していたのはわずか35%であった。このことから、腸内細菌の大部分において、シキミ経路は転写的に不活性であり、これらの細菌は芳香族アミノ酸オーソトロフであることが示唆された。これはおそらく、腸内細菌が食餌からこれらの芳香族アミノ酸を獲得する能力があるためであろう。
グリホサートがヒトの健康に及ぼす可能性のある影響
多くのヒトを対象とした研究で、グリホサート暴露と生殖系、引用43 中枢神経系、引用44 免疫系の様々な障害との関連性が検討されている。引用45 グリホサート暴露と非ホジキンリンパ腫の発症との関連性が複数の研究で報告されている。同様に、グリホサートへの暴露は喘息、引用50自閉症スペクトラム障害、パーキンソン病との関連が指摘されている。
上記のヒトを対象とした研究の情報は、アンケートおよび/または電話インタビューによって収集された。このため、参加者が主観的にグリホサート暴露を報告するという点でバイアスがかかる余地がある。さらに、参加者は、除草剤の散布と除草剤を使用した家族の近くにいるという両方の点で、ラウンドアップTMへの曝露に基づいてグリホサート・リスクの程度を判断していた。グリホサートはラウンドアップTMの有効成分であるが、市販品には界面活性剤を含む他の化学物質が多数含まれており、それら自体がグリホサートとは無関係に害を引き起こす可能性がある。グリホサートへの暴露濃度を明確にしたヒトでの研究は、我々の知る限り存在しない。一つの例外はZollerらCitation54による研究で、既知濃度のグリホサート(196.8μg)とアミノメチルホスホン酸(AMPA)(1.67μg)を「試験皿」に入れて参加者に与えた。その後、尿中のグリホサートとAMPA濃度を調べた。しかし、このような状況であっても、参加者の食事中のグリホサートの存在にはばらつきがあるため、グリホサートとAMPAの濃度を決定することは難しい。動物モデルが必ずしもヒトでの研究結果を予測するとは限らないが、有用な情報源にはなりうる。動物モデルを用いて、グリホサートと肥満、糖尿病、自閉症、精神疾患などの発症との関連性が示されている。しかし、これらの研究では、選択した動物モデルの腸内細菌叢に対するグリホサートの影響は調べていない。したがって、これらの例では、腸内細菌叢の変化が疾患の発現に影響を及ぼす要因であるかどうかを判断することはできない。ヒトの腸内細菌叢と動物モデルの腸内細菌叢の違い、そしてそのために動物モデルで得られた結果がヒトでの結果を反映しない可能性があることにも触れておく価値がある。
図2. 動物実験に基づく、グリホサート暴露が様々な身体系に及ぼす可能性のある影響。Biorenderで作成。
図2. 動物実験に基づく、グリホサート暴露が様々な身体系に及ぼす可能性のある影響。Biorenderを使用して作成。
フルサイズ表示
宿主が利用するトリプトファンの大部分は食事から摂取されるが、腸内細菌もまた、宿主が利用するトリプトファンの産生に関与している。グリホサートは微生物が産生するトリプトファンを減少させることが示されていることから、グリホサートは宿主におけるトリプトファンの利用可能性を減少させることが示唆される。トリプトファンとその誘導体は、腸内細菌叢と宿主細胞の両方をつなぐシグナル伝達分子としての役割が研究されており、その結果、ヒトの健康や疾病に影響を及ぼしている。引用65 IBD患者の血清中のトリプトファン濃度の低下と、IBD症状の重症度上昇との関連を示す証拠がある。 引用67 トリプトファン誘導体のその他の特性には、抗炎症作用、引用68 免疫反応の亢進、バリア機能の亢進などがある。引用69,引用70 これらの疾病結果はグリホサート暴露と直接関連づけることはできないが、グリホサート暴露と微生物産生トリプトファンの減少を関連づける文献が大量にあることから、その可能性が示唆される。
グリホサートと腸内マイクロバイオーム
先に述べたように、グリホサートは広範囲に汚染されているため環境中に存在している。引用71 サウスカロライナ州で実施された研究では、農家の90%の尿中に最大233ppbの濃度でグリホサートが検出された。 引用72 アメリカでは、一般市民の60~95%の尿中に2~3μg/Lの濃度でグリホサートが存在し、ヨーロッパ人の40~50%の尿中濃度は1μg/L未満である。 引用71 幼児と乳児の腎機能に対するグリホサートの影響を調べた別の研究では、調査した新生児の30%の尿中に1.06μg/L未満の濃度でグリホサートが存在することが示された。 引用73 これは、母乳や粉ミルクにグリホサートが含まれていたため、あるいはグリホサートが胎盤を通過する能力のために起こった可能性がある。上記の研究で報告されたグリホサート濃度はすべて、EU(EFSA)の許容一日摂取量(ADI)である0.5 mg/kg BW/日の5%を超えていない(引用13)。
腸内細菌叢の構成と機能性に対するグリホサートの影響
グリホサートは、図3に示すように2つの異なる方法で分解される。第一の経路ではサルコシンが蓄積され、その後脱リン酸化によってグリホサートのC-P結合が切断され、グリシンが生成されるCitation76。第二の経路では、酵素グリホサート酸化還元酵素を介してC-N結合が切断され、グリオキシレートとAMPAが生成されるCitation77。 引用77 自然界には、アルスロバクター属、GLP-1、Geobacillus caldoxylosilyticus T20、シュードモナス属、リゾビウム属のような多数の細菌を含む、様々な生物がグリホサートを分解することができる、 Citation78,Citation79自然界で特定の細菌がグリホサートを分解する能力を持つことを考慮すると、腸内細菌叢の細菌がグリホサートを分解する可能性が仮定されている。ある研究では、純粋なグリホサート(シグマアルドリッチ社製)を15人の参加者の糞便懸濁液と嫌気的にインキュベートすることにより、ヒトの糞便微生物叢におけるグリホサートの分解を調べた。このことから、ヒトの腸内細菌叢はグリホサートを異化できず、その結果グリホサートは排泄されるため、この化合物の生物濃縮の可能性は限定的であることが示唆される。サルコシン、グリシン、グリオキシル酸は尿中に存在することが示されている。引用81-84 しかし、これはグリホサートの分解を検討した研究の一部として実証されたものではないため、グリホサートの代謝物が体外にどのように排泄されるかについての結論は得られない。ある研究では、最初に摂取された全グリホサート(グリホサート、CAS Bioflow® 1071-83-6)の1%が尿中に排泄され、AMPA(AMPA、CAS Pro® 1066-51-9)の23%が尿中に排泄されることが明らかにされている。
図3. グリホサートの分解経路とそれに続く生成物。Biorenderを用いて作成。
図3. グリホサートの分解経路とその結果生じる後続生成物。Biorenderで作成。
フルサイズ表示
これらの代謝物の宿主への影響に関する情報はやや限られており、腸内細菌叢への影響はなおさらである。しかし、純粋なグリホサート(Interchim SS-7701)とその代謝物であるAMPA(Sigma-Aldrich 324,817)の宿主への影響を比較した研究が行われており、AMPAは腸内細菌叢に大きな影響を与えないことが示されている。この研究では、グリホサートがS. alviを無処置の対照群よりも5~13倍減少させることが示されている。引用85 尿サンプル中のAMPAの存在は乳がん患者とも関連しており、ある研究では、乳がん群では健常対照群と比較してAMPAを含むサンプルが38%増加したことが報告されている。 引用81 慢性腎臓病モデルラットでは、慢性腎臓病を代表する未治療群でグリシン共役化合物の血清レベルが上昇しており、グリシンの高レベルが慢性腎臓病と関連していることが示唆された。この現象はまた、Clostridium、Enterorhabdus、Parasutterella、Blautia、Escherichia Shigellaの増加と一致することが指摘されている。 引用86 ある研究では、グリオキシル酸の存在は宿主にとって有益であり、筋形成を促進し、筋萎縮を抑え、その後筋肉細胞でアミノ酸に代謝されることが示されている。引用87 サルコシンの増加は前立腺がんの発症と関連しており、引用88 サルコシンを病気のバイオマーカーとして利用する診断テストが数多く研究されているほどである。 引用89 対照的に、サルコシンは統合失調症の治療にも使用されており、抗精神病薬と併用することで顕著な効果が得られている。引用90 この分野はさらに研究する価値があるだろう。というのも、これらの代謝産物が宿主に及ぼす影響について概説した情報はあるが、腸内細菌叢に及ぼす影響や関連する生物学的効果に関する情報は乏しく、検討が必要だからである。
糞便のpHの上昇は、結腸内のグリホサート(バイオフロー®)の量と強い相関関係がある。引用91 同様に、グリホサート濃度の上昇は盲腸内の酢酸濃度に負の影響を及ぼす。グリホサートはトリプシン、リパーゼ、アミラーゼなどの消化酵素にも影響を及ぼすことが示されている。引用92 グリホサートは脂肪を消化するリパーゼとタンパク質を消化するトリプシンを阻害するという仮説がある。このため、無傷のタンパク質が大腸に入り、そこで分解されてアンモニアが放出され、大腸のpHが上昇する。
ビフィズス菌はヒトの腸内細菌叢の重要なメンバーとして認識されており、多くの菌株が健康促進作用を持つことが証明されている。 Citation94 ビフィズス菌の中には、母乳に含まれるムチンやヒトミルクオリゴ糖などの難消化性糖質を代謝し、宿主に重要な生理機能を発揮する短鎖脂肪酸(SCFA)を産生するものもあるCitation95 しかし、ビフィズス菌はグリホサートに対して最も感受性の高い細菌の一つであることが知られているCitation96 さらに、ほとんどの乳児用ミルクがグリホサートで汚染されていることが報告されている。ある研究では、0.03 mg kg-1から1.08 mg kg-1のレベルが報告されている。引用97 このことは、乳児の腸内におけるビフィズス菌減少の問題をさらに悪化させる可能性がある。
動物実験に基づくグリホサート関連マイクロバイオームの変化が健康に及ぼす可能性のある影響
近年、動物を用いた様々な研究が、図4に見られるように、腸内マイクロバイオームに対するグリホサートの影響を調べている。しかし、これらの研究で生物学的影響をもたらしたグリホサートの濃度は、ADIの3.5倍から500倍であることに注意すべきである。これらの研究により、グリホサートが関連する腸内細菌叢の変化が、体内の様々なシステムの障害と関連していることが明らかになった。具体的には、グリホサート(GLY、N-(phosphonomethyl)-glycine、純度95%以上)を含む飼料を250 mg/kg BWで2ヵ月間摂取した雄ラットでは、精子の運動性の低下、精子の奇形の増加、精巣構造の障害が認められた。ラットの腸内細菌叢の16S rRNA配列決定により、常在細菌集団に有意な変化が認められた。精子の質が低下したラットでは、プレボテラとバクテロイデスの増加が観察された。これらの知見は、グリホサートが誘発する腸内細菌異常症が雄性不妊症につながる可能性があるという仮説を支持するものである。
図4. 腸内細菌叢に対するグリホサートの影響と存在細菌種の増減を調べるために使用した動物モデルの例。 * 使用したグリホサートの濃度と実験期間。Biorenderを用いて作成。
図4. 腸内細菌叢に対するグリホサートの影響と存在細菌種の増減を調べるために使用した動物モデルの例。 * 使用したグリホサートの濃度と実験期間。Biorenderを用いて作成。
フルサイズ表示
ラットの盲腸内細菌叢は、グリホサート曝露(0.5、5、50 mg/kg体重/日)によって変化することが報告されている(ピュア・グリホサート、ラウンドアップTMバイオフロー®、レンジャー・プロ®)。この場合、バクテロイーダは減少し、バシロータと放線菌は増加した。Citation100 グリホサート暴露後の腸内細菌叢組成の変化に加えて、腸の完全性の変化も成体雄マウスで報告されている。これは、シンデカン-1プロテオグリカンの循環の変化と、ZO-1およびZO-2タイトジャンクションエフェクタータンパク質の発現によって確認された。この研究では、グリホサート(ラウンドアップTM)を妊娠中から成体まで、飲料水中に0.075%w/vの濃度で暴露した結果、バクテロイーダータ、シュードモナドータ、デスルホバクタータの相対レベルが変化した。
ホモシステインは、特定のビタミンB群の存在下でシステインに変換される代謝産物である。ビタミンB12、B6、葉酸の欠乏は、高ホモシステイン血症(血中に15μmol/Lを超えるホモシステインが存在する状態)を誘発することが示されている。純粋なグリホサートとラウンドアップ(Roundup®)をオスの子犬に投与(1.75 mg/kg bw/日、49日間)した場合の影響を調べたある研究では、この化合物が高ホモシステイン血症を誘発することが明らかになった。これは前述のLiuらの報告とは相反するものである。Citation99ではグリホサート(GLY、N-(ホスホノメチル)-グリシン、純度95%以上)処理によってプレボテラのレベルが上昇した。この結果の違いは、投与量の変更に起因している可能性があり、先に述べた研究と比較して、グリホサートの投与量がかなり少なかったためにプレボテラの量が減少したのである。脳障害、記憶や認知機能の低下、自閉症などの健康問題は、ホモシステインの増加と関連している。Citation60,Citation102 したがって、グリホサートへの早期暴露が、このような障害の発症の一因になっている可能性があると推測される。
グリホサートはまた、自閉症や脳症などの神経発達障害とも大きな相関関係がある(Barnettらによる総説)。これは、腸-脳軸全体の変化による可能性がある。グリホサートがルミノコッカス属、ビフィドバクテリウム属、ラクトバチルス属などの常在菌の生育を低下させる能力は、おそらく微生物代謝産物レベルの低下につながる。神経伝達物質として働くL-グルタミン酸や神経調節に関与するSCFAを含むこれらの代謝産物は、腸脳軸を通過する。L-グルタミン酸が減少することの意義は、GABAを合成する際の前駆体であることであり、GABAの減少は自閉症などの精神神経疾患と関連している。前述したように、シュードモナス属、アルスロバクター属、ジオバチルス属など、グリホサートに対して耐性を持つ細菌がいる。 これらの細菌は活性酸素種や炎症性サイトカインのレベルを上昇させ、視床下部-下垂体-副腎皮質(HPA)の活性化やグルココルチコイドの増加を引き起こし、神経発達に影響を及ぼす可能性がある。 引用106 自閉症の子どもは、そうでない子どもに比べてクロストリジウムの濃度が高いことが示されている。引用107 脳症とクロストリジウムが産生する毒性代謝産物との関連も明らかになっている。この相関関係から、グリホサートの腸内細菌叢への作用が、脳障害の一因となる毒性代謝産物の蓄積を引き起こす変化につながっているという説が提唱されている。
腸内マイクロバイオームに対するグリホサートの影響を調べるために、ミツバチやミミズを含む他の様々な動物モデルが一般的に用いられてきた。3種のミミズについて、ラウンドアップ®(グリホサート7.20 g/l)を115.49 mL/m2の濃度で2日間連続暴露した後の腸内細菌群集の乱れを調べた。引用109 グリホサートのミツバチへの影響は、ミツバチが受粉において重要な役割を担っていることから、多くの研究テーマとなっている。亜致死量の純粋なグリホサート(10 mg/L)(有効成分、シグマ・アルドリッチ社製)に慢性的に曝露したミツバチが、ノセマ・セラナエ(成虫に見られる病原性細菌)に7日間感染した結果、免疫反応が変化し、感染に抵抗できなくなった。 引用110 実際、グリホサート曝露とノセマ・セラナエ感染後、ミツバチのマイクロバイオームの組成が変化した。特に、Lactobacillus apisとSnodgrassella alviの存在量が減少した。これらの細菌種がミツバチの腸内マイクロバイオームの中心メンバーであることを考えると、これは重要な観察結果である。別の研究では、0.1 µg/L(3日間)の濃度でミツバチに対する純粋なグリホサートの影響を調べた。これは蜂蜜に自然に含まれる濃度を反映したものである。この場合、ミツバチのマイクロバイオームは非中核細菌種のみの減少を示した。Citation111 この研究では、前述の研究に比べてグリホサートの濃度を大幅に下げたことに注意すべきである。
腸内マイクロバイオームに対するグリホサートの影響を調べるために、高度な分子プロファイリングを利用した研究はわずかである。この知識のギャップに対処するため、Mesnage ら Citation112 は、グリホサートとグリホサートの液体製剤である MON 52276 の両方を、1 日あたり 0.5, 50, 175 mg/kg 体重(BW)の濃度で 90 日間曝露したラットの盲腸サンプルについて、メタボロームおよびメタゲノム配列決定を行いました。予想通り、メタボローム研究により、ラットの盲腸マイクロバイオームにおけるシキミ酸経路がグリホサートによって阻害されることが確認された。 引用100 腸内マイクロバイオームの多様性に関しては、グリホサート処理後にAcinetobacter johnsoniiとEggerthella属が増加した。アシネトバクターの増加が健康に及ぼす影響はまだ不明であるが、Eggerthella lentaのようなEggerthella属の中には、消化管感染症に関連しているものもある。 Citation114 ラウンドアップTMの超低用量(50 ng/L)を毎日暴露したラットを調べた同じグループによる、2年以上にわたる先行研究がある。Citation115 この場合、ラットは非アルコール性脂肪性肝疾患を発症しており、低レベルのグリホサートが肝臓に悪影響を及ぼすという仮説をさらに裏付けている。
ルーメンモデルもまた、グリホサートが細菌群集に与える影響の程度を調べるために利用されてきた。Ackermannら(引用116)によって実施されたそのような研究のひとつは、最終濃度0、1、10、100μg/mlのグリホサート(N-ホスホノメチル)グリシンをルーメン液発酵モデルの処理に用いたものである。グリホサートはまた、微生物集団の特定のグループの減少に関連し、一方で特定の潜在的病原性細菌を増加させたが、これらの結果は有意ではなかった。Riede ら(Citation117)は、PlantacleanR 360(1リットルあたりグリホサートイソプロピルアミン塩 486 g)を配合した RUSITEC ルーメン液発酵で、グリホサートによる変化が最小限であることを観察した。同様に、細菌群集のわずかな変動が観察されたが、グリホサート暴露が有益な常在細菌を特異的に標的としたり、病原性細菌の増殖を促進したりする兆候は認められなかった。Nielsen ら(Citation91)は、Bioflow® 450 PLUS(グリホサート酸当量 450 g/L)5~80mg/mLがヒツジのルーメン発酵モデルに及ぼす影響を調べた。これらの結果も、最小限の抗菌活性が観察されたという点で、前述の研究と一致していた。ただし、この結果は芳香族アミノ酸が十分なレベルにある環境に依存していた。
グリホサート処理作物の摂取が腸内細菌叢に及ぼす影響
前述したように、農作物や家畜飼料にグリホサートが残留していることは広く報告されている。過去数年間、グリホサートは主に作物の播種前の雑草防除手段として使用されていたが、ラウンドアップTM対応作物の開発により、除草剤を成長中の植物に直接散布できるようになったため、一年中グリホサートを使用できるようになった。この開発により、人間が消費する作物や家畜の飼料用作物にグリホサートが残留する可能性が高まった。モンサント社は、グリホサートの哺乳類に対するリスクは最小であり、グリホサートが残留している飼料や作物は心配ないと報告している。引用118 グリホサートで汚染された飼料(73.8および84.5 mg/日)を16週間摂取した牛のルーミナルマイクロバイオームを調査したある研究では、ルーミナルマイクロバイオームの構成に悪影響はなかったと判定されている。飼料の調製には、収穫前のグリホサート処理が含まれた。
結論
1974年の発売以来、グリホサートはますます環境の一部となり、特に1996年にはラウンドアップTM耐性作物が発売された。その結果、飲料水や農作物、汚染された空気の吸入を通じて、人間や家畜がグリホサートを摂取する機会が増えた。ラウンドアップTMの広範な使用は、人間の生理学のほとんどすべての側面に影響を及ぼす様々な障害の増加と相関している。これは相関関係であり、因果関係ではないことを認識することが重要である。グリホサート暴露の悪影響を強調する研究が増えている以外には、グリホサートの流通とヒトの障害の有病率の増加とを明確に関連付けた研究は、私たちの知る限り存在しない。
これまでの動物実験では、グリホサートが体内のさまざまな重要なシステムに影響を及ぼす可能性があることが説得力を持って示唆されている。しかし、ヒトにおける研究はやや矛盾しており、グリホサートとガン発生、喘息の重症度上昇、関節炎との関連を示す研究もあれば、グリホサートはこれらの障害に影響を与えないとする研究もある。我々の知る限り、グリホサートと人体との相互作用を調査したすべてのヒト研究は、アンケートや電話インタビューによるグリホサート暴露の自己申告のみに依存している。このような主観的な情報収集手段を考慮すると、ヒトを対象とした研究においてグリホサート暴露について得られるデータはやや限られている。動物モデルにおけるグリホサート暴露に関する知見は豊富にある。これらの研究は、EFSAが安全と判断したADI(0.5 mg/kg BW/日)でグリホサートを投与すると害が生じることを示している。このことは、ADI以下のグリホサートに暴露されたラットで腎臓と肝臓の機能障害が観察されたSéraliniらによる引用120でも確認されている。しかし、これらの研究がヒトにどのように反映されるかは不明である。
腸内細菌叢に関しては、グリホサート暴露後の変化として、バクテロイデス属菌の増加、ビフィドバクテリウム属菌、ルミノコッカス属菌、ラクトバチルス属菌の減少が挙げられる。腸内細菌叢のこうした変化は、高ホモシステイン血症、生殖毒性、腸の完全性の変化、神経発達障害、毒性代謝物の蓄積などの生物学的影響と関連している。しかし、これらの研究で用いられたグリホサートの濃度はADIを上回っていた。EFSAが決定したADIを用いてラットを処理した試験の一例では、Eggerthella、A. johnsonii、A. muciniphilaの増加が報告されている。このことは、ADIで投与されたグリホサートは、少なくともこの試験の正確な条件下では、生物学的影響をもたらすほど重大なマイクロバイオーム障害を引き起こさないようであることを示唆している。この濃度で腸内細菌叢に有意な障害がないのは、これらの細菌がグリホサートに対して抵抗性を示すためである可能性が高い。グリホサートに対する腸内細菌の感受性または耐性に関する報告は、存在するEPSPS遺伝子のクラスに焦点を当てた場合、やや矛盾しているが、シキミ経路が多数の腸内細菌で転写的に作動していないことを示唆するメタデータの解析は、より説得力がある。これは、腸内環境が細菌に十分なレベルの芳香族アミノ酸を供給しているため、腸内細菌叢がこれらの芳香族アミノ酸を産生する必要性が最小限に抑えられているためと考えられる。前述したように、食事は宿主の細胞機能だけでなく、宿主に存在する細菌の機能にも必要な芳香族アミノ酸を供給することができるが、摂取した食事がグリホサート暴露の結果、すでに芳香族アミノ酸が枯渇していなければの話である。
EFSAが決定したグリホサートのADIは、動物モデルにおいて害を引き起こすのに十分な高濃度であるが、これが腸内細菌叢の乱れによるものであるとは考えにくい。しかし、ヒトに関するグリホサート研究には大きな隔たりがある。既知の濃度のグリホサートで治療する対照試験が必要であろう。倫理の観点からは実現不可能かもしれないが、そのような研究が不足しているため、グリホサートが腸内細菌叢とヒトの健康全般の両方に及ぼす影響を完全に解明することは困難である。EFSAは2023年12月以降、グリホサートのEUでの使用を認可しないと発表した。それにもかかわらず、ヒトにおけるグリホサートレベルと腸内マイクロバイオームのシグネチャーとの関連は定義されるべきである。最後に、グリホサートへの曝露がマイクロバイオームの機能に及ぼす影響についての知識が不可欠である。
謝辞
本書は、アイルランド科学財団(SFI)の助成番号SFI/12/RC/2273_P2(APCフェーズ2センター助成番号APC18753-テーマ1.1 Microbes to Molecules)の資金援助を受けて実施された研究から生まれた。欧州連合(ERC、BACtheWINNER、プロジェクト番号101054719)の助成を受けた。ただし、記載された見解および意見は著者個人のものであり、必ずしも欧州連合または欧州研究会議執行機関の見解を反映するものではない。欧州連合および助成機関はそれらに責任を負うことはできない。
情報開示
著者による潜在的な利益相反は報告されていない。
その他の情報
資金提供
本研究はアイルランド科学財団(SFI/12/RC/2273_P2)の助成を受けた。欧州連合(ERC、BACtheWINNER、プロジェクト番号101054719)の助成を受けた。ただし、記載された見解および意見は著者個人のものであり、必ずしも欧州連合または欧州研究会議執行機関の見解を反映するものではない。欧州連合(EU)および欧州研究会議執行機関は、これらについて責任を負うことはできない。
参考文献
ベンブルックCM. 米国および世界におけるグリホサート除草剤使用の傾向。Environ Sci Europe. 2016;28(1):1–16. doi:10.1186/s12302-016-0070-0. [Crossref] [PubMed], [Google Scholar].
Chaturvedi P, Shukla P, Giri BS, Chowdhary P, Chandra R, Gupta P, Pandey A. Prevalence and hazardous impact of pharmaceutical and personal care products and antibiotics in environment: 新興汚染物質に関するレビュー。環境中の医薬品、パーソナルケア製品、抗生物質の有病率と有害影響:新興汚染物質のレビュー。 [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
Glyphosate: response to comments, usage and benefits. Office of Chemical Safety and Pollution Prevention; 2019. [Google Scholar].
Duke SO. 化学農薬およびその製剤成分と植物および植物害虫に関連する微生物との相互作用。J Agr Food Chem. 2018;66(29):7553–7561. doi:10.1021/acs.jafc.8b02316. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
グリホサート除草剤の高感度検出のための分子インプリントポリマーベースの電気化学センサー。Int J Environ Anal Chem. 2015;95(15):1489–1501. doi:10.1080/03067319.2015.1114109. [Taylor & Francis Online] [Web of Science ®], [Google Scholar].
Majewski MS, Coupe RH, Foreman WT, Capel PD. ミシシッピ州の大気と雨に含まれる農薬:1995年と2007年の比較。Environ Toxicol Chem. 2014;33(6):1283–1293. doi:10.1002/etc.2550. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
高等植物におけるケイ素を介した生物ストレス緩和のメカニズム:総説。Environ Pollut. 2007;147(2):422–428. doi:10.1016/j.envpol.2006.06.008. [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
Chang F, Simcik MF, Capel PD. 除草剤グリホサートとその分解物アミノメチルホスホン酸の大気中における存在と運命。Environ Toxicol Chem. 2011;30(3):548–555. doi:10.1002/etc.431. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
除草剤グリホサートの微生物群集への影響を通じた植物、動物、ヒトの健康への間接的影響。この論文では、除草剤グリホサートの微生物群集への影響を通して、植物や動物、ヒトの健康に間接的な影響を及ぼすことを明らかにした。 [Crossref] [Web of Science ®], [Google Scholar].
ウィリアムズGM、アーデマM、アッカヴェッラJ、ベリーSC、ブルシックD、バーンズMM、デ・カマルゴJLV、ガラブラントD、グリームHA、キアーLD. 独立した4つの専門家パネルによるグリホサートの発がん性の検討とIARC評価との比較。Crit Rev Toxicol. 2016;46(sup1):3–20. doi:10.1080/10408444.2016.1214677. [Taylor & Francis Online] [PubMed] [Web of Science ®], [Google Scholar].
EFSA. グリホサートの内分泌かく乱作用の可能性に関する農薬リスクアセスメントの査読。EFSA J. 2017;15:e04979. [PubMed] [Web of Science ®], [Google Scholar].
Rueda-Ruzafa L, Cruz F, Roman P, Cardona D. Gut microbiota and neurological effects of glyphosate. Neurotoxicology. 2019;75:1–8. doi:10.1016/j.neuro.2019.08.006. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
EFSA. 活性物質グリホサートの農薬リスク評価のピアレビューに関する結論。EFSA J. 2015;13:4302. [Web of Science ®], [Google Scholar].
Belkaid Y, Harrison OJ. 恒常性免疫と微生物叢。Immunity. 2017;46(4):562–576. doi:10.1016/j.immuni.2017.04.008. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Heiss CN, Olofsson LE. 腸内細菌叢に依存したエネルギー代謝の調節。J Innate Immun. 2018;10(3):163–171. doi:10.1159/000481519. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
腸内細菌叢のコロニー形成抵抗性と腸内病原体感染のメカニズム。Front Cell Infect Microbiol. 2021;11:1273. doi:10.3389/fcimb.2021.716299. [Crossref] [Web of Science ®], [Google Scholar].
Patterson E, Cryan JF, Fitzgerald GF, Ross RP, Dinan TG, Stanton C. Gut microbiota, the pharmabiotics they produce and host health. Proc Nutr Soc. 2014;73(4):477-489. doi:10.1017/S0029665114001426. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
O'Sullivan JN, Rea MC, Hill C, Ross RP. 皮膚微生物叢を操作する生物学的ツール。FEMS Microbiol Ecol. 2020;96(6):fiaa085. doi:10.1093/femsec/fiaa085. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
Dang AT, Marsland BJ. 微生物、代謝産物、腸-肺軸。Mucosal Immunol. 2019;12(4):843–850. doi:10.1038/s41385-019-0160-6. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Cryan JF, O'Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, Codagnone MG, Cussotto S, Fulling C, Golubeva AV. 微生物叢-腸-脳軸。Physiol Rev. 2019;99(4):1877-2013. doi:10.1152/physrev.00018.2018. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Cénit MC, Matzaraki V, Tigchelaar EF, Zhernakova A. Rapidly expanding knowledge on the role of the gut microbiome in health and disease. Biochim Biophys Acta - Mol Basis Dis. 2014;1842(10):1981–1992. doi:10.1016/j.bbadis.2014.05.023. [Crossref] [Web of Science ®], [Google Scholar].
Carding S, Verbeke K, Vipond DT, Corfe BM, Owen LJ. 疾患における腸内細菌叢のディスバイオシス。Microb Ecol Health Dis. 2015;26(1):26191. doi:10.3402/mehd.v26.26191. [テイラー&フランシスオンライン] [PubMed]、[Googleスカラー]。
腸内細菌叢組成は高齢者の食事と健康と相関する。Nature. 2012;488(7410):178–184. doi:10.1038/nature11319. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Erlandson KM, Liu J, Johnson R, Dillon S, Jankowski CM, Kroehl M, Robertson CE, Frank DN, Tuncil Y, Higgins J. 運動介入は、高齢の座りがちな成人の便微生物叢と代謝物を変化させる。Ther Adv Infect Dis. 2021;8:20499361211027068. doi:10.1177/20499361211027067. [Crossref] [Web of Science ®], [Google Scholar].
Durack J, Lynch SV. 腸内細菌叢:疾患との関係と治療の機会。J Exp Med. 2019;216(1):20–40. doi:10.1084/jem.20180448. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Clemente JC, Manasson J, Scher JU. 全身性炎症性疾患における腸内細菌叢の役割。Bmj. 2018;360:j5145. doi:10.1136/bmj.j5145. [Crossref] [PubMed], [Google Scholar].
Mirza A, Mao-Draayer Y. The gut microbiome and microbial translocation in multiple sclerosis. Cl Immunol. 2017;183:213–224. doi:10.1016/j.clim.2017.03.001. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Lopez MJ, Mohiuddin SS (2020). 生化学、必須アミノ酸。 [Google Scholar].
Zobiole LHS, Bonini EA, de Oliveira RS, Kremer RJ, Ferrarese-Filho O. Glyphosate affects lignin content and amino acid production in glyphosate-resistant soybean. Acta Physiol Plant. 2010;32(5):831–837. doi:10.1007/s11738-010-0467-0. [Crossref] [Web of Science ®], [Google Scholar].
Carbonari CA, Gomes GLGC, Velini ED, Machado RF, Simões PS, de Castro Macedo G. Glyphosate effects on sugarcane metabolism and growth. Amer J Plant Sci. 2014;5(24):3585. doi:10.4236/ajps.2014.524374. [Crossref], [Google Scholar].
コロンビアで栽培されたコカ植物におけるアルカロイド生成に対するグリホサートの見かけ上の影響。また、「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」「科学的根拠がない」。
Klebsiella pneumoniaeの5-enolpyruvylshikimate-3-phosphate synthase. 2. グリホサート[N-(phosphononmethyl)glycine]による阻害。Eur J Biochem. 1984;143(2):351-357。doi:10.1111/j.1432-1033.1984.tb08379.x. [Crossref] [PubMed], [Google Scholar].
除草剤に対する生物の感受性を評価するためのグリホサート標的酵素(5-enolpyruvylshikimate-3-phosphate synthase)の分類。J Hazard Mater. 2021;408:124556. doi:10.1016/j.jhazmat.2020.124556. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
[Googleスカラー] ゲインズTA、デュークSO、モランS、リゴンCAG、トラネルPJ、キュッパーA、ダヤンFE. 進化した除草剤耐性のメカニズム。J Biol Chem. 2020;295(30):10307–10330. doi:10.1074/jbc.REV120.013572. [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
Gomes MP, Juneau P. グリホサートによって誘発されるカモガヤ(Lemna minor L.)の酸化ストレス:ミトコンドリア電子伝達鎖は除草剤の標的か?Environ Pollut. 2016;218:402–409. doi:10.1016/j.envpol.2016.07.019. [クロスリフ] [PubMed] [Web of Science ®], [Google Scholar].
Staub JM, Brand L, Tran M, Kong Y, Rogers SG. 大腸菌膜排出トランスポーターの過剰発現によってもたらされる細菌グリホサート耐性。J Ind Microbiol Biotechnol. 2012;39(4):641–647. doi:10.1007/s10295-011-1057-x. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
グリホサートはヒトの微生物叢に影響するか?生命。2022;12(5):707. doi:10.3390/life12050707. [Crossref] [PubMed], [Google Scholar].
大腸菌分離株に対するグリホサートおよびグリホサート含有除草剤製剤の最小発育阻止濃度-病原性分離株と非病原性分離株および宿主種間の差異-. Front Microbiol. 2019;10:932. doi:10.3389/fmicb.2019.00932. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T. A human gut microbial gene catalog established by metagenomic sequencing. Nature. 2010;464(7285):59–65. doi:10.1038/nature08821. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
Rajilić-Stojanović M, Biagi E, Heilig HGHJ, Kajander K, Kekkonen RA, Tims S, de Vos WM. 過敏性腸症候群患者の糞便サンプルにおける微生物叢シグネチャーのグローバルかつ深い分子解析。Gastroenterology. 2011;141(5):1792–1801. doi:10.1053/j.gastro.2011.07.043. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
グリホサートを代謝するPseudomonas sp. Micro. 1988;54(12):2953–2958. [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
Mesnage R, Antoniou MN. グリホサートがヒト腸内細菌叢のシキミ酸経路に及ぼす影響について、計算モデリングにより明らかにした。Curr Res Toxicol. 2020;1:25–33. doi:10.1016/j.crtox.2020.04.001. [Crossref] [PubMed], [Google Scholar].
Arbuckle TE, Lin Z, Mery LS. オンタリオ州の農家集団における農薬曝露が自然流産のリスクに及ぼす影響の探索的分析。Environ Health Perspect. 2001;109(8):851–857. doi:10.1289/ehp.01109851. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
ワシントン州における農薬への推定住宅暴露とパーキンソン病による早期死亡率。Int J Env Res Pub He. 2018;15(12):2885. doi:10.3390/ijerph15122885. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
de Roos AJ, Cooper GS, Alavanja MC, Sandler DP. 農業健康調査における女性の関節リウマチ:農作業と曝露に関連するリスク。Ann Epidemiol. 2005;15(10):762–770. doi:10.1016/j.annepidem.2005.08.001. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
農薬とその他の農業リスク要因。アイオワ州およびミネソタ州の男性における非ホジキンリンパ腫の農薬およびその他の農業危険因子。Cancer Res. [PubMed] [Web of Science ®], [Google Scholar].
Eriksson M, Hardell L, Carlberg M, Åkerman M. 非ホジキンリンパ腫の危険因子としての農薬暴露(病理組織学的サブグループ解析を含む)。Int J Cancer. 2008;123(7):1657–1663. doi:10.1002/ijc.23589. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
非ホジキンリンパ腫と農薬曝露の症例対照研究。Cancer. 1999;85(6):1353–1360. doi:10.1002/(SICI)1097-0142(19990315)85:6<1353:AID-CNCR19>3.0.CO;2-1. [Crossref][PubMed][ウェブオブサイエンス®],[Google Scholar].
非ホジキンリンパ腫、非ホジキンリンパ腫、非ホジキンリンパ腫、非ホジキンリンパ腫、非ホジキンリンパ腫、非ホジキンリンパ腫。男性における非ホジキンリンパ腫と特定の農薬曝露:農薬と健康に関するカナダ横断研究。Cancer Epidemiol Biomarkers Prev. 2001;10:1155-1163. [PubMed] [Web of Science ®], [Google Scholar].
Hoppin JA, Umbach DM, London SJ, Henneberger PK, Kullman GJ, Alavanja MCR, Sandler DP. 農業健康調査における農家女性の農薬とアトピー性喘息および非アトピー性喘息。Am J Respir Crit Care Med. 2008;177(1):11–18. doi:10.1164/rccm.200706-821OC. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
von Ehrenstein OS, Ling C, Cui X, Cockburn M, Park AS, Yu F, Wu J, Ritz B. Prenatal and infant exposure to ambient pesticides and autism spectrum disorder in children: population based case-control study. Bmj. 2019;364:l962. doi:10.1136/bmj.l962. [Crossref] [PubMed], [Google Scholar].
Henneberger PK, Liang X, London SJ, Umbach DM, Sandler DP, Hoppin JA. 喘息を持つ農薬散布者における症状の増悪。Int Arch Occup Environ Health. 2014;87(4):423–432. doi:10.1007/s00420-013-0881-x. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
カメルF、タナーCM、アンバックDM、ホッピンJA、アラバンジャMC、ブレアA、コミンズK、ゴールドマンSM、コレルM、ラングストンJW。農業健康調査における農薬曝露と自己申告によるパーキンソン病。Am. J. Epidemiol. 2006;165(4):364–374. doi:10.1093/aje/kwk024. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
経口暴露の定量的バイオマーカーとしての尿中グリホサート濃度。Int J Hyg Envir Heal. 2020;228:113526. doi:10.1016/j.ijheh.2020.113526. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
De Long NE, Holloway AC. 早期の化学物質曝露とメタボリックシンドロームのリスク。DMSO. 2017;Volume 10:101-109. doi:10.2147/DMSO.S95296. [Taylor & Francis Online], [Google Scholar].
Prasad M, Gatasheh MK, Alshuniaber MA, Krishnamoorthy R, Rajagopal P, Krishnamoorthy K, Periyasamy V, Veeraraghavan VP, Jayaraman S. Impact of glyphosate on the development of insulin resistance in experimental diabetic rats: role of NFκB signalling pathways. 抗酸化物質。2022;11(12):2436. doi:10.3390/antiox11122436. [Crossref] [Web of Science ®], [Google Scholar].
Beecham JE, Seneff S. 自閉症とグリホサート系除草剤との間に関連性はあるか?J Autism. 2016;3(1):1. doi:10.7243/2054-992X-3-1. [Crossref], [Google Scholar].
Aitbali Y, Ba-M'hamed S, Elhidar N, Nafis A, Soraa N, Bennis M. グリホサート系除草剤への曝露は、マウスの腸内細菌叢、不安、うつ様行動に影響を及ぼす。Neurotoxicol Teratol. 2018;67:44–49. doi:10.1016/j.ntt.2018.04.002. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
遺伝子組み換え作物、腸内細菌叢、シキミ酸経路とシトクロム調節異常。Nutr Perspect J Council On Nutr. [Googleスカラー]。
Hu J, Lesseur C, Miao Y, Manservisi F, Panzacchi S, Mandrioli D, Belpoggi F, Chen J, Petrick L. グリホサート系除草剤の低用量曝露は、尿メタボロームと腸内細菌叢との相互作用を破壊する。この論文では、グリホサート系除草剤の低用量曝露が、尿メタボロームと腸内細菌叢との相互作用を破壊することを明らかにした。 [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
Drašar P、Poc P、Stárka L.グリホサート、重要な内分泌撹乱物質。Diabetol Metab Endokrinol Vyziv。2018;47:777-780. [Google Scholar].
Dai P, Hu P, Tang J, Li Y, Li C. Effect of glyphosate on reproductive organs in male rat. Acta Histochem. 2016;118(5):519–526. doi:10.1016/j.acthis.2016.05.009. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
ガレゴスCE、バイアーCJ、バルトスM、ブラスC、ドミンゲスS、モナコN、グミラールF、ギメネスMS、ミネッティA. ラットにおける周産期のグリホサート系除草剤曝露は、脳の抗酸化状態、グルタミン酸およびアセチルコリン代謝を変化させ、認識記憶に影響を及ぼす。Neurotox Res. 2018;34(3):363-374. doi:10.1007/s12640-018-9894-2. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Kreutz LC, Barcellos LJG, de Faria Valle S, de Oliveira Silva T, Anziliero D, dos Santos ED, Pivato M, Zanatta R. 亜致死濃度のグリホサートに短期暴露したシルバーキャットフィッシュ(rhamdia quelen)における血液学的および免疫学的パラメータの変化。Fish Shellfish Immunol. 2011;30(1):51–57. doi:10.1016/j.fsi.2010.09.012. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
Roager HM, Licht TR. 健康および疾患における微生物トリプトファン異化物。Nat Commun. 2018;9(1):3294. doi:10.1038/s41467-018-05470-4. [Crossref] [PubMed], [Google Scholar].
Nikolaus S, Schulte B, Al-Massad N, Thieme F, Schulte DM, Bethge J, Rehman A, Tran F, Aden K, Häsler R. トリプトファン代謝の増加は炎症性腸疾患の活動性と関連している。Gastroenterology. 2017;153(6):1504–1516. doi:10.1053/j.gastro.2017.08.028. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Rothhammer V, Mascanfroni ID, Bunse L, Takenaka MC, Kenison JE, Mayo L, Chao C-C, Patel B, Yan R, Blain M. Type I interferons and microbial metabolites of tripleptophan modulate astrocyte activity and central nerve system inflammation via the aryl hydrocarbon receptor. Nat Med. 2016;22(6):586–597. doi:10.1038/nm.4106. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Schirmer M, Smeekens SP, Vlamakis H, Jaeger M, Oosting M, Franzosa EA, Ter Horst R, Jansen T, Jacobs L, Bonder MJ. ヒト腸内細菌叢と炎症性サイトカイン産生能との関連。Cell. 2016;167(4):1125–1136. doi:10.1016/j.cell.2016.10.020. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
種間・細胞間シグナル伝達分子としてのインドールの役割。Trends Microbiol. 2015;23(11):707–718. doi:10.1016/j.tim.2015.08.001. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Roager HM, Hansen L, Bahl MI, Frandsen HL, Carvalho V, Gøbel RJ, Dalgaard MD, Plichta DR, Sparholt MH, Vestergaard H. Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat microbiol. 2016;1(9):1–9. doi:10.1038/nmicrobiol.2016.93. [Crossref] [Web of Science ®], [Google Scholar].
Gillezeau C, van Gerwen M, Shaffer RM, Rana I, Zhang L, Sheppard L, Taioli E. The evidence of human exposure to glyphosate: a review. Environ Health-Glob. 2019;18(1):1–14. doi:10.1186/s12940-018-0435-5. [Crossref] [PubMed], [Google Scholar].
農業農薬の研究における曝露の誤分類:バイオモニタリングからの洞察。Epidemiology. 2006;17(1):69–74. doi:10.1097/01.ede.0000190603.52867.22. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
乳幼児におけるグリホサート暴露と腎障害バイオマーカー。環境汚染。2020;256:113334. doi:10.1016/j.envpol.2019.113334. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
グリホサートとアミン酸の分析。栄養成分および牛乳中のグリホサートおよびアミノメチルホスホン酸の、フルオレニルメチルオキシカルボニルクロライドによる誘導体化と液体クロマトグラフィー質量分析による分析。J Agr Food Chem. 2015;63(48):10562–10568. doi:10.1021/acs.jafc.5b04453. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
カナダ、ケベック州東部タウンシップにおける遺伝子組み換え食品に関連した農薬への母体および胎児の暴露。2011;31(4):528-533. doi:10.1016/j.reprotox.2011.02.004. [Crossref], [Google Scholar].
Dick RE, Quinn JP. 環境試料から分離されたグリホサート分解菌:発生と分解経路。Appl Microbiol Biotechnol. 1995;43(3):545–550. doi:10.1007/BF00218464. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
LBr株におけるグリホサートの代謝。1988;54(12):2953–2958. doi:10.1128/aem.54.12.2953-2958.1988. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
有機リン化合物の微生物分解。また、そのような微生物は、有機リン化合物の分解を促進することが示唆されている。
また、そのような環境下での農薬の使用は、農薬の安全性を確保する上でも重要であり、そのような環境下での農薬の使用は、農薬の安全性を確保する上でも重要である。Int J Env Res Pub He. 2020;17(20):7519. doi:10.3390/ijerph17207519. [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
Huch M, Stoll DA, Kulling SE, Soukup ST. ヒト糞便微生物叢によるグリホサートの代謝。(1)グリホサートはヒトの糞便微生物によって代謝される。 [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
Franke AA, Li X, Shvetsov YB, Lai JF. グリホサート代謝物アミノメチルホスホン酸の尿中排泄と乳がんリスクに関するパイロットスタディ:多民族コホート研究。Environ Pollut. 2021;277:116848. doi:10.1016/j.envpol.2021.116848. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Jentzmik F, Stephan C, Miller K, Schrader M, Erbersdobler A, Kristiansen G, Lein M, Jung K. 直腸指診後の尿中のサルコシンは、前立腺癌の検出および侵攻性腫瘍の同定におけるマーカーとしては不向きである。Eur Urol. 2010;58(1):12–18. doi:10.1016/j.eururo.2010.01.035. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Jones AR. 実験動物によるグリシン結合体の尿中排泄に関するいくつかの観察。Xenobiotica. 1982;12(6):387–395. doi:10.3109/00498258209052480. [テーラー&フランシスオンライン] [PubMed] [Web of Science ®], [Google Scholar].
林ELC、マトックスJK、ダニエルFB. ジクロロ酢酸の雄性Fischer 344ラットにおける組織分布、排泄および尿中代謝産物。J Toxicol Environ Health, Part A Curr Issues. 1993;38(1):19–32. doi:10.1080/15287399309531697. [Taylor & Francis Online] [PubMed] [Web of Science ®], [Google Scholar].
グリホサートは、その代謝物AMPAではなく、ミツバチの腸内細菌叢を変化させる。PloS One. 2019;14(4):e0215466. doi:10.1371/journal.pone.0215466. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
馮Y-L, 曹G, 陳D-Q, Vaziri ND, 陳L, 張J, 王M, 郭Y, 趙Y-Y. Microbiome-metabolomics reveals gut microbiota associated with glycine-conjugated metabolites and polyamine metabolism in chronic kidney disease. Cell Mol Life Sci. 2019;76(24):4961-4978. doi:10.1007/s00018-019-03155-9. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
グリシン由来のα-ケト酸代謝産物であるグリオキシル酸はC2C12細胞の筋形成を促進する。栄養素。2023;15(7):1763. doi:10.3390/nu15071763. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Sreekumar A, Poisson LM, Rajendiran TM, Khan AP, Cao Q, Yu J, Laxman B, Mehra R, Lonigro RJ, Li Y. メタボローム・プロファイルから、前立腺癌の進行におけるサルコシンの潜在的役割が明らかになった。Nature. 2009;457(7231):910–914. doi:10.1038/nature07762. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Mbage B, Li Y, Si H, Zhang X, Li Y, Wang X, Salah A, Zhang K. 葉酸官能基化ポリオキソメタレートナノ粒子の作製による比色測定におけるH2O2とサルコシンの同時検出。Sens Actuators B Chem. 2020;304:127429. doi:10.1016/j.snb.2019.127429. [Crossref] [Web of Science ®], [Google Scholar].
Lane H-Y, Liu Y-C, Huang C-L, Chang Y-C, Liau C-H, Perng C-H, Tsai GE. 急性統合失調症に対するサルコシン(N-メチルグリシン)治療:無作為化二重盲検試験。Biol Psychiatry. 2008;63(1):9–12. doi:10.1016/j.biopsych.2007.04.038. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
ニールセンLN、ローガーHM、カサスME、フランセンHL、ゴーゼウィンケルU、ベスターK、リヒトTR、ヘンドリクセンNB、バールMI。グリホサートは、十分な芳香族アミノ酸レベルにより、腸内環境における常在細菌群集組成への短期的影響は限定的である。Environ Pollut. 2018;233:364–376. doi:10.1016/j.envpol.2017.10.016. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
楊X、宋Y、張C、龐Y、宋X、呉M、程Y. グリホサート系除草剤ラウンドアップの中国産ミナミイチョウガニ(eriocheir sinensis)の生存、免疫反応、消化活動および腸内細菌叢に及ぼす影響。Aquat Toxicol. 2019;214:105243. doi:10.1016/j.aquatox.2019.105243. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Hidalgo-Cantabrana C, Delgado S, Ruiz L, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and their health-promoting effects. 薬としてのバグ: Ther Microbes Prev Treat Dis. 2018;79:7628-7638. [Google Scholar].
Saturio López S, Nogacka A, Alvarado-Jasso GM, Salazar N, González de Los Reyes-Gavilán C, Gueimonde Fernández M, Arboleya S. Role of bifidobacteria on infant health. 微生物。2021;9(12):2415. doi:10.3390/microorganisms9122415. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
ビフィズス菌と免疫系との分子コミュニケーション. Front Microbiol. 2017;8:2345. doi:10.3389/fmicb.2017.02345. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
The effect of glyphosate on potential pathogens and beneficial members of poultry microbiota in vitro. Curr Microbiol. 2013;66(4):350–358. doi:10.1007/s00284-012-0277-2. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
Rodrigues NR, de Souza APF. ブラジルで販売されている大豆ベースの乳児用調製粉乳中のグリホサートおよびAMPA残留物の発生。Food Addit Contam. 2018;35(4):724–731. doi:10.1080/19440049.2017.1419286. [Taylor & Francis Online] [Web of Science ®], [Google Scholar].
Walker M. 母乳栄養児への粉ミルク補給:有用か危険か?ICAN:乳幼児、児童、思春期栄養。2015;7(4):198-207。 [Crossref], [Google Scholar].
グリホサートが誘発する腸内細菌叢異常は、ラットの雄性生殖毒性を促進する。Sci Total Environ. 2022;805:150368. doi:10.1016/j.scitotenv.2021.150368. [クロスリフ] [PubMed] [Web of Science ®], [Google Scholar].
グリホサートとその製剤。グリホサートとその製剤であるラウンドアップ・バイオフローおよびレンジャープロは、ラットの盲腸マイクロバイオームにおける細菌および真菌の群集組成を変化させる。 [Google Scholar]
Del Castilo I, Neumann AS, Lemos FS, de Bastiani MA, Oliveira FL, Zimmer ER, Rêgo AM, Hardoim CCP, Antunes LCM, Lara FA. グリホサート系除草剤ラウンドアップ®の低用量終生暴露は、マウスに腸管障害、腸内細菌異常症、行動変化を引き起こす。この論文では、グリホサート系除草剤ラウンドアップ(RoundUp®)の低用量長期暴露による腸管障害と腸内細菌異常症、およびマウスの行動変化について報告する。 [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
血漿ホモシステイン値の上昇:認知症とアルツハイマー病発症の危険因子か危険マーカーか?J Alzheimer's Dis. 2006;9(4):393–398. doi:10.3233/JAD-2006-9404. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
バーネットJA、バンディML、ギブソンDL. 現代農業におけるグリホサートの使用は、腸-脳-マイクロバイオーム軸の調節を通じて精神神経症状の増加をもたらすか?フロント。2022;9. doi:10.3389/fnut.2022.827384. [Crossref] [Web of Science ®], [Google Scholar].
スペンサーCM、Alekseyenko O、ハミルトンSM、トーマスAM、Serysheva E、Yuva-ペイラーLA、ペイラーR.Fmr1KOマウスの行動表現型を変更する:遺伝的背景の違いは、自閉症様の反応を明らかにする。2011;4(1):40-56. doi:10.1002/aur.168. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
食餌性グルタミン酸による腸脳軸の活性化とエネルギー恒常性における生理的意義。また、そのような食事療法を行うことで、より効果的な食事療法を行うことが可能となる。 [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
脳における酸化ストレス:神経変性疾患における生存を支配する新規細胞標的. Prog Neurobiol. 2005;75(3):207–246. doi:10.1016/j.pneurobio.2005.02.004. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
を用いた、泌尿器科医による泌尿器科疾患に関する研究。自閉症スペクトラム障害児におけるクロストリジウム菌の影響と身体計測。J Mol Neurosci. 2020;70(6):897–907. doi:10.1007/s12031-020-01482-2. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Zuo Z, Fan H, Tang X, Chen Y, Xun L, Li Y, Song Z, Zhai H. 最小肝性脳症患者における異なる治療法とアルコール中毒が腸内細菌叢に及ぼす影響。Exp Ther Med. 2017;14(5):4887–4895. doi:10.3892/etm.2017.5141. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
グリホサート農薬の農地への影響。グリホサート系除草剤が3種のミミズ(alma millsoni、eudrilus eugeniae、libyodrilus violaceus)の腸内マイクロバイオームに及ぼす影響:パイロット研究。(1)ミミズの腸内細菌叢がミミズの腸内細菌叢に及ぼす影響. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
グリホサートの亜致死量への慢性曝露がミツバチの免疫、腸内細菌叢、病原体による感染に及ぼす影響。Microorganisms. 2021;9(4):845. doi:10.3390/microorganisms9040845. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Almasri H, Liberti J, Brunet J-L, Engel P, Belzunces LP. 農薬への軽度な慢性曝露は、中核的な腸内細菌叢を乱すことなく、ミツバチの健康の生理学的マーカーを変化させる。(1):1-15。doi:10.1038/s41598-022-08009-2。 [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Mesnage、Teixeira、Mandrioli、Falcioni、Ducarmon、QR、Zwittink、RD、Mazzacuva、F、Caldwell、Halket、J、Amiel、C. ショットガンメタゲノミクスとメタボロミクスを用いた、グリホサートまたはラウンドアップ MON 52276 の Sprague-Dawley ラットの腸内細菌叢と血清メタボロームへの影響評価。Environ Health Perspect. 2021;129(1):017005. doi:10.1289/EHP6990. [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
(注1)本論文は、本邦における臨床的・微生物学的特徴について検討したものである。卵菌血症の臨床的および微生物学的特徴。J Clin Microbiol. 2015;53(2):626–635. doi:10.1128/JCM.02926-14. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Nayfach S, Shi ZJ, Seshadri R, Pollard KS, Kyrpides NC. ヒト腸内細菌叢の未培養ゲノムから得られた新たな知見。Nature. 2019;568(7753):505–510. doi:10.1038/s41586-019-1058-x. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Mesnage R, Antoniou MN. グリホサートの毒性に関する議論における事実と誤り。Front Public Health. 2017;5:316. doi:10.3389/fpubh.2017.00316. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Ackermann W, Coenen M, Schrödl W, Shehata AA, Krüger M. The influence of glyphosate on the microbiota and production of botulinum neurotoxin during ruminal fermentation. Curr Microbiol. 2015;70(3):374–382. doi:10.1007/s00284-014-0732-3. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Riede S, Toboldt A, Breves G, Metzner M, Köhler B, Bräunig J, Schafft H, Lahrssen-Wiederholt M, Niemann L. "rumen simulation technique "による、グリホサート含有除草剤がin vitroのルーメン代謝と細菌に及ぼす可能性のある影響の調査。J Appl Microbiol. 2016;121(3):644–656. doi:10.1111/jam.13190. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Sørensen MT, Poulsen HD, Katholm CL, Højberg O. グリホサートの飼料残留-家畜の健康と生産性への潜在的影響。Animal. 2021;15(1):100026. doi:10.1016/j.animal.2020.100026. [クロスレフ] [PubMed] [Web of Science ®], [Google Scholar].
Billenkamp F, Schnabel K, Hüther L, Frahm J, von Soosten D, Meyer U, Höper D, Beer M, Seyboldt C, Neubauer H. No hints at glyphosate-induced ruminal dysbiosis in cows. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
Séralini G-E, Clair E, Mesnage R, Gress S, Defarge N, Malatesta M, Hennequin D, de Vendômois JS. ラウンドアップ除草剤とラウンドアップ耐性遺伝子組換えトウモロコシの長期毒性。Environ Sci Europe. 2014;26(1):1–17. doi:10.1186/s12302-014-0014-5. [Crossref], [Google Scholar].
PDFをダウンロード
関連研究
おすすめ記事
こんな記事も読まれています
引用元
情報提供者
著者
研究開発関係者
編集者
図書館員
学会
オープンアクセス
概要
オープンジャーナル
オープンセレクト
Dove Medical Press
F1000Research
機会
別刷りとe-prints
広告ソリューション
早期出版
企業アクセスソリューション
ヘルプと情報
ヘルプ&コンタクト
ニュースルーム
すべてのジャーナル
書籍
最新情報
ご登録いただくと、個別のリサーチやリソースをEメールでお届けします。
登録する
テイラー・アンド・フランシス・グループのFacebookページテイラー・アンド・フランシス・グループのTwitterページテイラー・アンド・フランシス・グループのLinkedinページ
テイラー・アンド・フランシス・グループYoutubeページテイラー・アンド・フランシス・グループWeiboページ
著作権 © 2023 Informa UK Limited
プライバシーポリシー
クッキー
利用規約
アクセシビリティ
イングランド&ウェールズ登録第3099067号
5 ハウィック・プレイス|ロンドン|SW1P 1WG
テイラー・アンド・フランシス・グループ
AcceptCookieポリシー
当社は、お客様のウェブサイト体験を向上させるためにクッキーを使用しています。当社のクッキーの使用およびクッキーの設定を管理する方法については、当社のクッキーポリシーをご覧ください。このメッセージを閉じることで、クッキーの使用に同意したことになります。
×
この記事が気に入ったらサポートをしてみませんか?