皮膚恒常性と免疫における肥満細胞-微生物叢クロストークの新たな役割

MDPIオープンアクセスジャーナル
zoom_out_map検索メニュー
雑誌 Cells 12巻 22号 10.3390/cells12222624
設定論文別刷り注文
オープンアクセス総説
皮膚恒常性と免疫における肥満細胞-微生物叢クロストークの新たな役割
https://pubmed.ncbi.nlm.nih.gov/37998359/

キャメロン・ジャクソン・ボスヴェルド1、コリン・グース1、ナタチット・リムジュニャウォン2、*ORCIDおよびプリアンカ・プンディール1,*著1ゲルフ大学生物科学部分子細胞生物学科、ゲルフ、ON N1G 2W1、カナダ2マヒドン大学シリラジ病院医学部研究部、アレルギー・免疫学研究センター、バンコク10700、タイ*著者宛先Cells 2023, 12(22), 2624; https://doi.org/10.3390/cells12222624受領済み: 受理:2023年10月16日 / 改訂:2023年11月7日 / 受理:2023年11月9日 / 掲載:2023年11月14日 2023年11月9日 / 掲載:2023年11月14日(この論文は特集号「マスト細胞生物学への新たな洞察」に属しています。)Downloadkeyboard_arrow_down 図の閲覧 レビュー レポート バージョン ノート要旨皮膚には多面的なマイクロバイオームが存在し、細菌、真菌、ウイルスがバランスよく共存している。これらの常在微生物は、有害な病原体に対抗すると同時に、皮膚の免疫と協調して働くことにより、皮膚の健康を維持する上で基本的な存在である。ディスバイオーシスとして知られるこのバランスの乱れは、乾癬やアトピー性皮膚炎のような疾患の原因となる。皮膚の防御システムの中心にあるのが肥満細胞である。マスト細胞は皮膚層内に戦略的に配置され、潜在的な外敵に対して迅速に反応できるよう準備されている。最近の研究では、これらの肥満細胞と皮膚のマイクロバイオーム内の多様な存在との間の複雑な相互作用が解明され始めている。この関係は、特にバランスが取れている時と取れていない時の両方において、以前認識されていた以上に、皮膚の健康にとって不可欠であることが証明されつつある。本総説では、マスト細胞と皮膚常在微生物の結びつきに関する最新の知見を紹介し、皮膚の健康と疾患に対するそれらの複合的な影響に光を当てる。キーワード:肥満細胞;マイクロバイオーム;皮膚免疫;ホメオスタシス;宿主-微生物
はじめに
皮膚は身体の中で最大の器官であり、多面的な役割を担っている。紫外線、温度、微生物、毒素、アレルゲンなど、多くの種類のダメージから身体を守っている[1,2]。皮膚には、ケラチノサイト、線維芽細胞、ランゲルハンス細胞(LC)、樹状細胞(DC)、マクロファージ、αβおよびγδT細胞、ナチュラルキラーT(NKT)細胞、マスト細胞など、多くの免疫細胞や常在細胞が関連している。これらの細胞はすべて、皮膚のホメオスタシスだけでなく、防御の第一線で異なる役割を果たしている [3,4]。また、皮膚表面には多くの常在細菌、真菌、ウイルスが生息しており、これらが一体となって皮膚マイクロバイオームを形成している [5] 。皮膚マイクロバイオームは、病原体に対する防御において重要な役割を担っており、この恒常性が乱れると、疾病が引き起こされる可能性がある [6] 。皮膚免疫とマイクロバイオータの相互作用の複雑さは、共生的な意味合いを持つが、今日の西洋的なライフスタイルにおいては、そこに存在する免疫細胞に対する皮膚マイクロバイオータの影響により、自己免疫疾患や炎症性疾患が増加しているため、今後も問題を引き起こす可能性がある [5] 。皮膚に存在する免疫細胞の中でも、マスト細胞は、炎症反応を引き起こし、他の自然免疫細胞や適応免疫細胞を動員するように設計された分泌顆粒で満たされた組織常在の自然免疫細胞である [4,7] 。本総説では、特にマスト細胞と皮膚常在菌との相互作用に焦点を当て、皮膚における免疫系とマイクロバイオームとの複雑な関係に光を当てる。皮膚自然免疫
皮膚の自然免疫細胞は、病原体に対する身体の防御と皮膚の健康維持に重要な役割を果たしている。皮膚内では、免疫細胞はそのほとんどが真皮に存在し、侵入してくる微生物に対する防御の第一線を担っている。皮膚免疫には複雑な組織と多様な免疫細胞集団が関与しており、詳細な議論については別の文献でレビューされている [8,9,10]。ここでは、免疫と炎症の最前線に位置する特殊な自然免疫細胞であるマスト細胞のみに焦点を当てる。皮膚には肥満細胞の割合が最も多く、マウスの耳の皮膚では白血球集団の10%を肥満細胞が占めている [7,11]。(a)肥満細胞は、血管や神経の近くに戦略的に位置することで、空間的な優位性を持っており、異物を素早く察知して反応することができる。(b)さらに、活性化後数秒以内に高密度に詰まった分泌顆粒を貯蔵・放出する能力を持つため、時間的優位性があり、必要な時に迅速かつ増幅された免疫応答が可能である [12]。全体として、肥満細胞は上皮と強い結びつきがあり、表皮のバリア機能と皮膚の恒常性の維持に重要な役割を果たしている [13,14] 。多くの研究が、バリア機能と免疫バランスを維持する上で、マスト細胞と皮膚の近傍の免疫細胞および非免疫細胞との間の細胞間コミュニケーションの重要性を強調している [4] 。その結果、不必要な免疫活性化から生じる病態の発症を避けるためには、皮膚コンパートメントのもう一つの常在細菌に対する身体の反応を厳密に制御することが極めて重要である。マスト細胞は、皮膚常在細菌を認識し許容する一方で、潜在的な病原体に対しては警戒を怠らず、微妙なバランスを保つのに役立っている。マスト細胞は免疫応答を調節し、免疫寛容を促進することで、常在細菌群との調和のとれた共存を保証することができる。本講演では、常在細菌と皮膚に常在する肥満細胞との間の複雑なつながりについてさらに考察し、肥満細胞の発達、機能、そして健康と疾病のバランスをとる行為への影響を解明する。皮膚における肥満細胞-概要
3.1. 肥満細胞のサブタイプと発生
すべての肥満細胞は、Paul Ehrlichによって最初に同定された陽イオン性メタクロマチック色素による独特の染色に基づき、他の白血球とは著しく異なっているが、それぞれの組織部位における異なる起源と組織微小環境に由来する複数の表現型のバリエーションを示し、その結果、異なる機能的特殊化が生じている。これらのサブセットは、異なるサイトカイン発現、顆粒内容、受容体プロファイルを示す(表1)。マウスとヒトの肥満細胞は、受容体の発現やその命名法も異なっている。マウスでは、成熟時の解剖学的位置に基づいて定義された2つの主要な肥満細胞サブクラスがあります:ほとんどの結合組織(例えば、皮膚、腹膜腔、気管、舌、食道など)に見られる結合組織肥満細胞(CTMC)と、主に呼吸器や腸の粘膜の内側に位置する粘膜肥満細胞(MMC)です。ヒトの場合、これらはトリプターゼ+キマーゼ+肥満細胞(MCTC)とトリプターゼ+肥満細胞(MCT)に相当する。その名が示すように、肥満細胞の表現型の最も明らかな違いは、トリプターゼとキマーゼの両方を産生するか、放出可能な顆粒内でトリプターゼのみを産生するかである。最近、Tauberらは、偏りのないシングルセルRNAシーケンス(scRNA-seq)を用いて、多臓器にわたるマウスとヒトの肥満細胞集団の分子的差異を解析し、ヒト肥満細胞の不均一性は、マウス肥満細胞のそれよりもはるかに複雑であることを発見した[15]。トランスクリプトーム・プロファイルに基づき、ヒトの様々な臓器に分布する7つの潜在的な肥満細胞サブセットが示され、そのうち3つの異なるサブセットは皮膚にのみ特異的に認められた。
表1. ヒトにおけるトリプターゼに富む肥満細胞とトリプターゼに富む肥満細胞の違い。
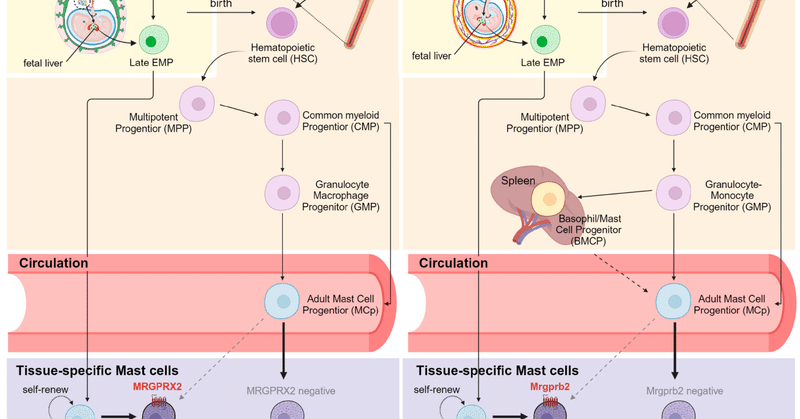
さらに、CTMCsとMMCはその発生起源において食い違いがあることが明らかである(図1)。CTMCsの大部分は、胎生期に胎児肝臓で移動・発達する卵黄嚢由来の赤血球・骨髄系前駆細胞(EMP)から生じ、構成的に存在し、長期間持続する、 MMCの寿命はわずか2週間と短く、骨髄由来の無顆粒球性造血幹細胞(HSC)前駆細胞と容易に置換可能である [7,16,17,18,19]。注目すべきは、成体マウスの皮膚におけるCTMCsは、MMCと比較して、ドナー由来の造血幹細胞の再構成がはるかに乏しいことが、いくつかの骨髄養子移植研究によって確認されていることで、皮膚において、長寿命で組織常在性の肥満細胞の前駆細胞が自己再生していることが示唆されている [18,20]。その起源に関わらず、肥満細胞の発生は、幹細胞因子(SCF)、形質転換成長因子-β、CCL2、IL-3、アクチビンなどの組織特異的な成長因子の影響を受ける[21,22]。
細胞 12 02624 g001図1. ヒトとマウスにおける肥満細胞の発生。(A)ヒトの組織特異的マスト細胞は、胎生期に子宮内で発生し、標的部位に移動し、組織内の前駆細胞を介して自己維持するか、あるいは出生後に骨髄で発生し、循環を通じてコミットしたマスト細胞前駆細胞を介して更新する。(B)マウスの肥満細胞の発生は、胚でも骨髄でも同じように始まる。マウス脾臓でも、好塩基球-肥満細胞前駆細胞(BMCP)が好塩基球と肥満細胞のいずれかに分化できることを示唆する証拠がいくつかある。図はBioRenderで作成。
3.2. 皮膚における肥満細胞の解剖学的位置
皮膚に存在する肥満細胞の多くは、皮膚の表皮層の下にあり、神経、血管、毛包、脂肪組織、筋肉組織の近くにある [3,21]。肥満細胞と神経線維の解剖学的関係は、様々な臓器で観察される。皮膚では、肥満細胞は、肥満細胞メディエーターであるヒスタミンの受容体を発現するC型およびA型末梢神経線維に近接して存在する。逆に、肥満細胞は古典的な神経伝達物質(アセチルコリンやコルチコトロピン放出ホルモンなど)や神経ペプチド(サブスタンスPやカルシトニン遺伝子関連ペプチドなど)に対する多数の受容体を発現している。この肥満細胞と神経の相互作用はフィードバックループを形成し、肥満細胞から放出されたヒスタミンが神経ペプチドの放出を誘発する。これらの神経ペプチドは、今度は肥満細胞の脱顆粒を刺激し、ヒスタミンとプロスタグランジンD2(PGD2)の放出を引き起こし、このサイクルを持続させる。
同様に、血管と皮膚肥満細胞との間の解剖学的な関連とその結果生じる相互作用は、多面的である。これらの相互作用には、血液内皮細胞(BECs) との結合や循環中の細胞との相互作用が含まれる [4,21]。皮膚肥満細胞からのヒスタミン放出は、血流の増加、血管内皮カドヘリン(CD144)の局在の変化、血管系の透過性亢進をもたらす [4,24]。マスト細胞はTNF⍺を血管壁を通して血流中に放出し、特に強固な接着と管腔内這い回りの際に、さらなる好中球をリクルートする [25] 。さらに、肥満細胞は複数の血管内皮増殖因子(VEGF-A、-B、-C、-D)を産生し、血管新生(VEGF-A、-B)とリンパ管新生(VEGF-C、-D)の両方に寄与する[4,25]。肥満細胞が血管と相互作用するもう一つの方法は、血管壁を通して糸状体を伸ばすことである。血管周囲肥満細胞はIgEを捕捉することができ、サンプリング機構として機能し、血管内を循環する際に他の細胞を勧誘する可能性がある [26,27]。皮膚肥満細胞はまた、ロイコトリエンを介して収縮を誘導することにより筋肉と相互作用し、免疫細胞のリクルーターおよびアクチベーターとしてインターロイキンを分泌し、炎症反応を誘発する [21] 。
真皮の肥満細胞はまた、樹状細胞(真皮DCおよびランゲルハンス細胞)、真皮マクロファージ、自然リンパ球(ILC)2、線維芽細胞、ケラチノサイト、メラノサイトなど、他の皮膚細胞と直接または間接的に接触している。ケラチノサイト、内皮細胞、平滑筋細胞、線維芽細胞、好酸球や成熟肥満細胞など、皮膚の構造的・免疫的細胞成分のいくつかは、肥満細胞の成熟、増殖、肥満細胞のアポトーシスの抑制、走化性、接着、脱顆粒の亢進を誘導する最も重要な因子の一つであるSCFの供給源であることが判明している [29,30] 。さらに、線維芽細胞との接触は、結合組織肥満細胞の表現型に関連する遺伝子の発現を増加させることが観察されている。その結果、肥満細胞はIL-4、IL-13、線維芽細胞増殖因子を分泌して線維芽細胞の増殖を促進したり [31,32] 、ヒスタミンを放出して線維芽細胞からのコラーゲン産生を開始したりする [33] 。皮膚マスト細胞はまた、接触依存的および接触非依存的な様式で、真皮DCと直接相互作用することが分かっている(文献[28]を参照)。さらに、肥満細胞は、メラノーマ細胞だけでなく、皮膚の特殊な色素産生細胞であるメラノサイトの増殖促進にも重要な役割を果たしていることを示す有力な証拠がある。この促進は、線維芽細胞増殖因子2(FGF-2)やIL-8のようなメディエーターの作用を通じて起こり、腫瘍の進行に寄与する [34,35] 。さらに、肥満細胞から放出されるヒスタミンは、H2受容体を介してメラノサイトのメラニン形成、遊走、形態に影響を及ぼす。紫外線もヒスタミン放出を誘発し、色素沈着を引き起こす可能性がある。メラノサイトおよび白斑性ケラチノサイトに対するこれらのヒスタミン作用は、白斑患者の再色素形成におけるヒスタミンの使用を支持するかもしれない [36,37,38]。
3.3. 皮膚肥満細胞の表現型的特徴
一般に、肥満細胞は独特であり、細胞質内に電子密度の高いリソソーム様分泌顆粒を多く含むことで容易に特徴付けることができる。これらの分泌顆粒には、様々なリソソーム酵素、肥満細胞特異的および肥満細胞非特異的プロテアーゼ、ヒスタミンやセロトニンなどの生体アミン、サイトカイン、成長因子など、多数の形成前メディエーターが含まれている [39] 。さらに、肥満細胞は、フローサイトメトリーを用いて、免疫グロブリンE(IgE)の高親和性レセプターであるFcεRI、SCFレセプターであるc-kit(CD117)といった細胞表面マーカーを共発現することにより、容易に同定することができる。同様に、その明確なトランスクリプトーム・プロファイルは、他の免疫細胞とも一線を画している [7] 。しかし、先に述べたように、マスト細胞には複数のサブセットがあり、それぞれがサイトカイン、ケモカイン、補体レセプターの産生を含む表現型や機能にバリエーションを示している。マウスやヒトに関するEliehら、Xingら、McNeilらの報告[40,41,42]に要約されている。皮膚肥満細胞に注目すると、他の結合組織肥満細胞、特に腹膜に見られる肥満細胞と多くの転写シグネチャーを共有している [7]。皮膚肥満細胞を含むこれらのCTMCsは、セリンプロテアーゼを担当する遺伝子(例えば、Ctsg(cat、 Ctsg(カテプシンGをコードする)、Mcpt2(肥満細胞プロテアーゼ2をコードする)、Mcpt4、Mcpt9、Tpsab1(トリプターゼα/β1をコードする)、Tpsb2、Tpsg1(トリプターゼγ1をコードする)、Cma1(キマーゼ1)、およびCma2)、ならびにエイコサノイド/プロスタグランジン生合成の重要な酵素であるプロスタグランジン合成酵素(例えば Hpgds(造血系プロスタグランジンD2合成酵素をコードする)など) [7]。これらのCTMCsのもう一つの重要な分子的特徴は、Mas関連Gタンパク質共役型受容体(MRGPR)のメンバーであるヒトMRGPRX2とそのマウスオルソログMrgprb2(図1)を独占的に発現していることである。MRGPRX2/Mrgprb2は、IgE/FcεRI非依存的なマスト細胞の脱顆粒に関与しており、神経ペプチドのような塩基性分泌促進分子やカチオン性ペプチドの幅広いレパートリーに応答するため、過去10年間で注目度が高まっている、 抗菌性宿主防御ペプチド(HDP)、化合物48/80(c48/80)、いくつかのFDA認可薬などの基本分泌分子やカチオン性ペプチドの幅広いレパートリーに反応して肥満細胞は脱顆粒し、生命を脅かす偽アレルギー反応を引き起こす可能性がある [41,44,45,46,47,48]。最近のscRNA-seq解析により、Mrgprb2の発現は皮膚肥満細胞を含むCTMCsに限定され、MrgprB2+対MrgprB2-肥満細胞は、マウスの異なる組織で一貫して保存されたトランスクリプトームコアを持つ、異なる肥満細胞サブセットを示すことが検証された [15]。
4. 皮膚微生物の多様性-健康な皮膚と炎症を起こした皮膚
4.1. 健康な皮膚マイクロバイオーム
皮膚微生物叢は、常在菌と呼ばれる細菌、真菌、ウイルスを含む多様な微生物から構成されている。間違いなく、細菌は様々な皮膚部位に分布する最も一般的な微生物である。16SリボソームRNA(rRNA)配列決定により、マイクロバイオーム常在細菌は通常3つの主な属に分類されることが明らかになっている: コリネバクテリア属、プロピオニバクテリア属、ブドウ球菌属である [49] 。これらは一般的に最も一般的な属であるが、特殊なニッチを形成する異なる環境の皮膚部位を比較すると、集団頻度はより変動する傾向がある。例えば、鼻の脇や頭皮の裏側など、皮脂が大量に分泌される部位では、親油性のプロピオニバクテリウム属が多くなる傾向がある一方、膝の裏などの湿った部位では、多量の汗を資源として利用するコリネバクテリア属やブドウ球菌属が多くなる [50] 。一方、真核生物リボソーム遺伝子の内部転写スペーサー1(ITS1)領域の塩基配列決定によって決定された真菌群集の構成は、異なる地形的皮膚部位に沿って比較的一貫している。マラセチア属真菌は、生理的属性に関係なく、ほとんどの部位で優勢な種であるが、足部など一部の部位では、より多様な真菌群集を示すこともある[51,52]。細菌や真菌とは異なり、ウイルス群集の多様性を調査することは、これらの微生物間で共有される普遍的なマーカー遺伝子が存在しないこと、真核生物のウイルスの存在は特定の解剖学的部位に関連するというよりもむしろ個体特異的であることが判明していることなどから、主にユニークな課題を提示している [51]。
健康な場合、皮膚マイクロバイオームはほぼ安定しているが、皮膚のpH、水分、生理学的組成の変化は、そこに存在する細菌に影響を及ぼす [51,53] 。このような違いには、例えば、乾燥、湿潤、皮脂分泌などの微小環境が含まれる [5,51]。多様性と群集の安定性は逆相関関係にあり、多様性が増すと安定性は低下する。健康な個体では、皮膚微生物叢は一般的に恒常的であり、もし群集が経時的に変化しても、種は類似のニッチと類似の数を持つ他の微生物に置き換わる可能性があり、その結果、集団全体としてはほとんど変化しない。皮膚マイクロバイオームには個人差があり、特に湿潤部位では、皮脂分泌部位と乾燥部位は個人間でより類似している。
微生物叢の常在菌は、病原体や疾患に対する防御をもたらす一方で、一部の微生物は、放置すると宿主に害を及ぼす可能性がある。これを防ぐために、ある種の生物は他の生物の増殖を制御して恒常性を維持することができる。例えば、黄色ブドウ球菌は日和見病原体であり、機会があればバイオフィルムを形成し、湿疹としても知られるアトピー性皮膚炎を引き起こす。これを防ぐために、常在菌である表皮ブドウ球菌は、Espとして知られるセリンプロテアーゼを分泌し、黄色ブドウ球菌がバイオフィルムを形成するために使用するタンパク質を分解することができる。
常在菌は、自らの手段で病原体の増殖を制御することに加え、常在する免疫細胞と相乗的に作用して免疫力を付与することができる。免疫反応を開始し強化することで知られる炎症性サイトカイン、インターフェロン-γ(IFN-γ)とインターロイキン(IL)-17Aの放出を、特異的病原体フリー(SPF)マウスと無菌(GF)マウスで比較した研究では、GFマウスのT細胞はSPFマウスに比べてIFN-γとIL-17Aの産生が有意に少なく、その結果、GFマウスでは他の病原体による感染を制御するのに不可欠な皮膚免疫が損なわれていることが判明した[55]。注目すべきことに、このT細胞サイトカイン産生能の低下は、主に皮膚微生物叢の欠如に起因するものであった。皮膚および皮膚排出リンパ節における他の免疫細胞の存在は、GFマウスとSPFマウスで同等であり、抗生物質の経口投与による微生物叢の減少(特に、皮膚ではなく腸内)は、皮膚T細胞によるこれらの炎症性サイトカインの産生に影響を及ぼさなかったからである。興味深いことに、S. epidermidisのような単一の皮膚常在菌を導入するだけで、皮膚におけるIL-17A産生を回復させる効果が証明された [55]。全体として、これらの知見は、常在細菌が皮膚内の免疫適性を維持するために不可欠であることを示している。マイクロバイオームと免疫反応の相乗効果を示すもう一つの例は、創傷治癒に見られる。創傷治癒プロセスにおいて、表皮菌が産生するリポテイコ酸は、ケラチノサイトからの過剰な炎症性シグナルの放出を防ぐことができるため、創傷治癒プロセスが制御され、免疫系が過剰に活性化するのを防ぐことができる [56]。
4.2. 疾患における皮膚マイクロバイオーム
アトピー性皮膚炎、アレルギー性接触皮膚炎、にきび、慢性創傷、免疫系の活性化による慢性炎症など、一般的な皮膚疾患の多くは、皮膚マイクロバイオームの変化と関連していることが示されている。健康な人は、環境暴露、宿主因子、および/または殺菌性産物の分泌により、皮膚マイクロバイオームの制御を失い、その結果、皮膚マイクロバイオームの多様性と安定性が低下することがある [51,53] 。例えば、アトピー性皮膚炎では、黄色ブドウ球菌の個体数が正常値を超えて増加し、これは疾患の再燃や症状の悪化と関連している [57] 。このような個体群のシフトが起こると、免疫系はその脅威を認識し、対応することができる。アトピー性皮膚炎における免疫系の反応のメカニズムのひとつは、黄色ブドウ球菌が産生するδ-毒素に反応して肥満細胞が脱顆粒し、自然免疫反応と適応免疫反応の両方が誘導されることである [24] 。ディスバイオーシスの役割は、他の疾患よりもアトピー性皮膚炎においてはるかに広く研究されているが、ディスバイオーシスと乾癬などの他の疾患との間に関連があることを示唆する証拠がある。乾癬患者では、健常人と比較して皮膚マイクロバイオーム集団に違いがあるが、この関係は決定的なものとは断定されていない [58] 。この関連を検証するためには、さらに調査が必要であり、またディスバイオーシスと他の皮膚疾患や感染症との関連についても調査が必要である。
皮膚微生物叢と免疫ネットワークは共に、病原体から防御し、皮膚の完全性を維持し、皮膚の免疫応答を制御する高度な防御システムを形成している。これらの構成要素間の相互作用を理解することは、皮膚の健康を促進し、皮膚関連疾患を治療するための効果的な戦略を開発するために不可欠である。
4.3. マスト細胞と常在細菌との相互作用
マスト細胞活性化の従来のよく知られた経路を図2に示す。マスト細胞が真皮に存在することは立証されているが [3,21]、細菌性病原体関連分子パターン(PAMPs) [2,3,59,60]と直接相互作用することから、細菌が皮膚の深層に侵入するメカニズムが存在することが示唆される。NakatsikiらとGriceらは、傷害部位のない正常なヒトの皮膚の皮下領域で常在細菌を検出したことから、細菌が皮膚の真皮層にまで侵入することを示した[49,61]。この論文では、16S rRNA遺伝子、特異的抗原、細菌rRNAをコードするDNAの存在のみを調査しているが、生きた細菌の存在を証明することはできず、その成分は、真皮に位置する肥満細胞やその他の免疫細胞が細菌のPAMPsによって直接活性化される可能性があることを示している。Bayらの研究でも、操作分類学的単位(OTU)の種の豊富さレベルは低いものの、皮膚細菌が健康な皮膚に浸潤する能力があることが示されている[62]。このことは、皮膚の真皮層では種の豊富さは減少するものの、マスト細胞や他の免疫細胞が、皮膚内の細菌と直接相互作用する可能性が残っていることを示している[62]。
細胞 12 02624 g002図2。異なる刺激による肥満細胞の活性化。(A)FcεR1を介したIgE介在性肥満細胞活性化。(B)PAMPsとDAMPsに反応するToll様受容体(TLRs)を介した非IgE介在性肥満細胞活性化。(C)補体を介した非IgE媒介性肥満細胞活性化。(D)MRGPRX2受容体を介した神経ペプチドおよびクオラムセンシング分子による活性化。BioRenderで作成した図。
5. 肥満細胞発生における皮膚マイクロバイオームの役割
肥満細胞の成熟における皮膚マイクロバイオームの役割は、最も重要である。この関係は、WangらによるGFマウスとSPFマウスの肥満細胞成熟を比較した研究で最も適切に示されている。この研究では、マスト細胞の成熟はSCFレセプターであるc-kitの高発現と定義され、未熟はc-kitの低発現と定義された。GFマウスはSPFマウスと比較して、皮膚中のSCF濃度が低下していることに加え、成熟肥満細胞が有意に少ないことがわかった [63]。さらに、成熟マスト細胞のこの減少は、GFマウスをSPFマウスに曝露して微生物叢を再構成すると回復した。この成熟の欠如をさらに強調するものとして、一般的な肥満細胞活性化物質である化合物48/80を注射することで誘発される後肢の炎症が、SPFマウスと比較してGFマウスでは減少していることがわかった[63]。この発見の背後にあるメカニズムを解明するために、Wangらは、ブドウ球菌のリポテイコ酸を注射すると、ケラチノサイトのSCF産生がアップレギュレートされ、その結果、GFマウスとSPFマウスの両方で、c-kit発現肥満細胞数を増加させることができることを示した。さらに、Cre-loxシステムを用いたScf遺伝子のケラチノサイト特異的ノックアウトは、これらの変異マウスの皮膚への肥満細胞の動員を完全に消失させ、肥満細胞の移動がケラチノサイトのSCF産生に完全に依存していることを明確に示した [63]。皮膚マイクロバイオームは、皮膚における肥満細胞のホーミングと成熟を制御する上で極めて重要な役割を果たしている。この制御は、スタフィロコッ クスのリポテイコ酸に反応するケラチノサイト由来のSCF産生 の調節を通して起こる(図3)。このことは、ケラチノサイトでSCFを過剰発現させると、皮膚の肥満細胞集団が増加するという観察によって、最初に証明された [63]。この研究に先駆けて、KunisadaらとHuttunenらによる研究が行われ、創傷治癒におけるケラチノサイト、肥満細胞、SCFの役割を明らかにしようとしたが、メカニズムの解明には至らなかった [64,65]。これらの研究は、皮膚マイクロバイオームと肥満細胞の成熟との関 係の背後にあるメカニズムについて有用な洞察を与えてくれるが、 この関係については、他にどのような細菌学的由来の分子がこのシス テムに影響を与えうるかなど、まだまだ発見すべきことが多い。
細胞 12 02624 g003図3. 皮膚常在細菌叢は肥満細胞の成熟を促進する。特定の病原体を持たない典型的なマウスでは、皮膚マイクロバイオーム由来のリポテイコ酸(LTA)の存在が、ケラチノサイトに十分な量の幹細胞因子(SCF)の産生を促し、これが肥満細胞の正しい成熟につながる。しかし、無菌マウスでは、SCFの供給が不十分なため、肥満細胞はc-Kitレセプターの発現低下を示す。図はBioRenderで作成。
6. 皮膚バリア機能における肥満細胞-無菌マウスと通常マウス
肥満細胞は表皮のバリア機能と創傷治癒を担っている [66,67,68]。マイクロバイオームは、皮膚バリアが破られたことを免疫系に知らせる上で極めて重要な役割を果たしている。通常の環境下で皮膚バリアが破られると、マイクロバイオームがPAMPsを放出し、マスト細胞などの常在免疫細胞によって検出される。マイクロバイオームが存在しない場合(GFマウスなど)、免疫系は当初、皮膚の表皮層にある傷害細胞からの危険関連分子パターン(DAMPs)によってのみ傷害部位に動員されることができる[66]。免疫系は微生物学的異物や病原体関連メディエーターによってプライミングされていないため、免疫系は未熟であると考えられる。その結果、皮膚バリアの創傷に対する初期反応が遅くなる [66]。マウスの正常な創傷治癒にはマスト細胞が必要であり、これはヒトにも当てはめることができる [68]。肥満細胞は、血管新生を促進するVEGF成長因子を大量に放出する。このVEGFの放出は、従来のSPFマウスよりもGFマウスの方が長く持続する [66]。肥満細胞はIL-10も産生し、他の肥満細胞との間でフィードバックループを作ることができる。このフィードバックはFcεRIレセプターのダウンレギュレーションを引き起こし、IgEの活性化を介して炎症因子の放出を抑制する。
7. 常在細菌に対する肥満細胞の耐性
マスト細胞は上皮と密接な関係を示し、バリア機能を支える役割を果たしている。従って、常在細菌との反応は、不当な免疫活性化の潜在的な結果を回避するために、細心の制御が必要である。最近、Di Nardoらは、肥満細胞と真皮線維芽細胞(dFB)との間に重要な関係があることを明らかにした[69]。彼らは、in vitroで肥満細胞とdFBを共培養し、その結果、肥満細胞が常在細菌に対して寛容になることを示した。dFBで調整したヒト肥満細胞は、IL-6、IL-8、Th2サイトカインIL-4、IL-5、IL-10、IL-13などの炎症性サイトカインの産生を抑制した。肥満細胞、常在細菌、dFB間の相互作用は互いに依存している。肥満細胞が成熟するにはマイクロバイオームが必要であり、マイクロバイオームが皮膚の肥満細胞を制御するにはdFBが必要なのである。そこで研究グループは、scRNA-seqを用いて、dFBとマスト細胞の共培養における違いを観察した。その結果、dFBにおける表現型の切り替えと、肥満細胞における免疫活性化遺伝子のダウンレギュレーションが観察された。NF-κBインヒビターα(NFKBIA)、すなわちA20/腫瘍壊死因子α誘導タンパク質3(TNFAIP3)もマスト細胞でアップレギュレートされた。RNA-seqとin silico解析により、CD44と細胞外マトリックス(ECM)との関連が指摘された。Toll様受容体-2(TLR2)はCD44の関連受容体であり、従って研究の次の段階のターゲットとなった。ヒアルロン酸(HA)は特定の組織においてTLR2のリガンドとして提案されており、ヒト肥満細胞におけるTLR2活性化の焦点であった。HA存在下ではTLR2は細胞表面から除去されるが、TLR4とCD44は発現されたままであることがわかった。dFBとの共培養はCD44を減少させた[69]。さらに研究者らは、線維芽細胞と肥満細胞間のコミュニケーションを仲介する重要なシグナル伝達経路、TGF-β経路を同定した。この経路の活性化は、肥満細胞の耐性を促進し、炎症反応を抑制する。この研究結果は、免疫恒常性の維持における皮膚細胞と肥満細胞との複雑な相互作用にさらに光を当てた [69]。要約すると、この研究ではECMの一成分であるHAが、TLR2のダウンレギュレーションを介してNF-κB経路を抑制することにより、常在細菌に対する肥満細胞の反応を抑制し、皮膚病原体に対する反応を低下させる経路を発見した(図4)。この研究はin vitroおよびex vivoの研究から得られた知見であるが、ヒトのin vivo実験での確認は不足している。この研究は、主に肥満細胞とdFBとの相互作用に焦点を当てており、肥満細胞の活性化に対する神経細胞のような他の細胞タイプの影響について、さらなる研究の必要性を示唆している。
細胞 12 02624 g004図4. 線維芽細胞はヒアルロン酸の産生を介して、常在細菌に対する肥満細胞の耐性を制御している。皮膚マスト細胞は真皮線維芽細胞との相互作用を通して常在細菌に対する耐性を示す。これらの線維芽細胞はNF-κBのA20阻害剤の発現を増加させることにより、肥満細胞の反応性を効果的に制御している。この複雑なメカニズムは、皮膚の恒常性を維持する一方で、他の外的課題に対する肥満細胞の反応を許容する役割を果たしている。図はBioRenderで作成。
同様に、上皮細胞や内皮細胞由来の産物は、皮膚微生物群に対する肥満細胞の耐性を調整する可能性がある。インターロイキン-33は、恒常性と炎症状態の両方において、上皮細胞、内皮細胞、線維芽細胞様細胞に豊富に発現する核内サイトカインである。細胞傷害や組織損傷時に放出され、ST2レセプターを発現する免疫細胞に注意を促す "アラミン "として働く [70] 。マスト細胞はIL-33の主要な標的の一つであり、両者の相互作用はアレルギー性疾患、感染性疾患、慢性炎症性疾患にとって極めて重要である。興味深いことに、恒常性においては、IL-33はマスト細胞を細菌の細胞壁成分であるリポ多糖(LPS)とペプチドグリカン(PGN)に対して無反応にする。LPSおよびPGNによって誘導される肥満細胞の活性化に対する抑制効果は、IL-33の濃度が最適値以下の場合に観察され、ST2経路を介して媒介される。ST2欠損動物由来のマスト細胞はLPSによって過剰に活性化されることから、IL-33はin vivoでの免疫恒常性維持過程においてマスト細胞の活性化を阻害することが示唆された。メカニズム的には、IL-33の濃度が最小になると、マスト細胞のインターロイキン-1受容体関連キナーゼ1(IRAK1)の分解が促され、LPSに反応しなくなり、常在細菌に対する免疫反応が回避される。逆に、感染や組織損傷の際には、IL-33レベルの上昇が肥満細胞を活性化し、炎症性サイトカインやケモカインの放出を促す [71] 。
常在細菌に対する肥満細胞の耐性の背後にあるメカニズムを理解することは、肥満細胞の活性化が重要な役割を果たしているアトピー性皮膚炎や乾癬などの炎症性皮膚疾患の治療法の開発につながる。例えば、Yuらによる研究では、皮膚常在菌が皮膚の構造と肥満細胞レベルに及ぼす影響について調査した。その結果、対照群の皮膚組織には真皮の異常がなく、正常で乱れのない構造を示した。逆に、アトピー性皮膚炎群では、表皮の肥厚、過角化症、有棘細胞症、表皮細胞の腫脹、真皮血管の拡張、炎症細胞の顕著な浸潤を特徴とする実質的な皮膚障害を示した。驚くべきことに、アトピー性皮膚炎グループの皮膚病変にS. epidermidisを塗布すると、マウスはコントロールグループと比較して皮膚障害が緩和された。トルイジンブルー染色で評価したところ、アトピー性皮膚炎群ではマスト細胞数が正常群に比べて有意に増加していた。注目すべきは、アトピー性皮膚炎+S.エピデルミディス群では、アトピー性皮膚炎群と比較して肥満細胞数が減少したことである。全体として、これらの結果は、皮膚常在菌がマウスの皮膚損傷を改善し、アトピー性皮膚炎の重症度を緩和する能力を有することを示唆している[60]。
いくつかの常在細菌は、TLR依存的経路とTLR非依存的経路の両方を介して、肥満細胞の脱顆粒を抑制する能力を実証している [72,73,74]。後者の経路では、大腸菌 [75]、乳酸桿菌 [76,77,78,79]、ビフィズス菌 [80]がFcεRIの細胞内シグナル伝達を阻害する。乳酸菌の多様な株は、マウスモデルにおいてアレルギー性皮膚炎を緩和する能力を示した [81,82,83,84,85,86,87]。さらに最近では、これらの知見がヒトの臨床試験にも応用され、小児および成人のアトピー性皮膚炎患者におけるプロバイオティクスの臨床的有効性が検討されている。プロバイオティクス(乳酸桿菌とビフィズス菌)のブレンドを6ヵ月にわたって定期的に摂取したところ、小児および青年のSCORingアトピー性皮膚炎(SCORAD)スコアが有意に低下したことが実証された [88] 。アトピー性皮膚炎の管理および予防における乳酸菌の利用可能性については、最近、Xieら(2023年)がまとめている [89]。
8. 常在細菌は病原体に対してマスト細胞を刺激する
皮膚微生物叢と肥満細胞との相互作用は、単なる発育と成熟にとどまらず、潜在的な病原体に対する肥満細胞の免疫機能の増強にまで及んでいる。その顕著な例が、皮膚常在菌である表皮ブドウ球菌由来のリポテイコ酸(LTA)が果たす役割であり、このLTAはマスト細胞の皮膚表面への動員を指揮する。皮膚で最も一般的なグラム陽性菌種である表皮ブドウ球菌は、TLR2のリガンドであるLTAを発現している。LTAを介してTLR2シグナルが活性化されると、マスト細胞はカテリシジン抗菌ペプチドの発現をアップレギュレートする。その結果、LTAで前処理した肥満細胞は、ワクシニアウイルス感染に対して高い抵抗性を示した。この抗ウイルス活性の高まりは、主にLTA誘導TLR2シグナル伝達に応答して肥満細胞が産生するカテリシジンに起因すると考えられる。その結果、皮膚表面にこれらの常在成分が存在することで、マスト細胞は潜在的な侵入病原体に対して常に準備態勢を整えていることになる。このことは、感染に対抗する能力を強化するだけでなく、身体の入口で用心深い歩哨として行動するよう、マスト細胞を準備させるのである [90]。
9. 細菌の種内および種間コミュニケーションは、肥満細胞を介した皮膚炎症に影響を与える。
細菌は他の種と情報を交換し、行動を調整するために様々なメカニズムを用いており、複雑なコミュニティを形成する能力を際立たせている。情報伝達の顕著な形態のひとつがクオラムセンシングであり、細菌はクオラムセンシング分子(QSM)と呼ばれる特定の化学シグナルを放出し、それを感知する。QSMは通常、グラム陰性菌ではアシルホモセリンラクトン、グラム陽性菌では自己誘導ペプチドなどの小さな分子である。これらの分子は、細菌集団が成長するにつれて蓄積し、細胞が自らの密度を測定することを可能にする。閾値濃度に達すると、バイオフィルム形成、病原性因子産生、遺伝子発現などの協調的な活動が開始される(総説[91,92,93])。マスト細胞は、受容体Mrgprb2(マウスのオルソログ)とMRGPRX2(ヒトのオルソログ)を介して、グラム陽性菌が産生するカチオン性QSMと相互作用することが報告されている [41]。その結果、肥満細胞の活性化とそれに続く脱顆粒が起こり、細菌が排除される(図5)。コンピテンス刺激ペプチド(CSP-1/-2)、エンテロバクチン合成酵素成分F(Entf)、ストレプチン-1などのQSMに加えて、MRGPRX2/b2は、Corbiereら[45,94]が要約したように、複数の抗菌ペプチドや宿主防御ペプチドとも相互作用することが報告されている。
細胞 12 02624 g005図5。マスト細胞はMRGPRレセプターを介して細菌間コミュニケーションを感知することができる。細菌は可溶性のクオラム感知分子(QSM)を産生し、その集団密度を知らせる。細菌集団が臨界量に達すると、これらのQSMは病原性に関連する特定の細菌遺伝子を活性化する。マスト細胞に特異的なレセプターであるヒトMRGPRX2とマウスMrgprb2は、グラム陽性菌由来のカチオン性細菌QSMを認識して結合することができる。この認識は肥満細胞の急速な脱顆粒を引き起こし、様々なメディエーター、すなわち抗菌性顆粒内容物やプロスタグランジンD2(PGD2)の放出につながる。この抗菌反応によって細菌が破壊され、抗菌特性を持つ他の免疫反応が開始される。BioRenderで作成した図。
興味深いことに、アトピー性皮膚炎の発症と制御に関してWilliamsらによって行われた研究から、ヒトの皮膚上の共生細菌と病原性細菌間のクオラムセンシングが、黄色ブドウ球菌によるダメージを抑制することで防御的役割を果たしていることが明らかになった。微生物によるディスバイオシスの間、黄色ブドウ球菌はフェノール可溶性モジュリン(PSM)αペプチドの活性を通して表皮バリアを障害する。これらのペプチドにさらされると、表皮の酵素活性が亢進し、皮膚バリアの破壊とそれに続く炎症を引き起こす。しかし、常在細菌叢、特にCoNS(コアグラーゼ陰性ブドウ球菌)と共存すると、黄色ブドウ球菌の皮膚に対する有害な影響は緩和される。これはクオラムセンシングAGRシステムの存在によるものである。agrシステムの構成成分であるI型自己誘導ペプチドは、黄色ブドウ球菌のagr活性を阻害し、黄色ブドウ球菌によって誘発される皮膚の炎症を緩和する。アトピー性皮膚炎の再燃時、ディスバイオーシスが深刻になると、これらの抑制性I型ペプチドの量が不足し、黄色ブドウ球菌が炎症に寄与するようになる。このことは、アトピー性皮膚炎に罹患した皮膚にPSMαが存在することからも裏付けられる。これらの知見は、皮膚マイクロバイオームの様々なメンバーが、黄色ブドウ球菌の疾患促進作用に対抗できる新たなメカニズムを明らかにするものである。このことは、黄色ブドウ球菌が過剰に存在し、細菌の多様性が低下することが、なぜアトピー性皮膚炎の症状の重症化につながるのかを説明できる可能性がある。アトピー性皮膚炎の病態生理におけるマスト細胞の極めて重要な役割と、細菌のクオラムセンシングコミュニケーションを遮断することでマスト細胞を宿主のために利用できる可能性を考慮すると、マスト細胞と皮膚マイクロバイオームの調和状態と不均衡状態の相互作用を理解することは、現在の治療アプローチを強化する道を開く可能性がある。
10. 結論
まとめると、本総説は、皮膚における肥満細胞と細菌微生物叢の相互作用に関する最新の理解を提供し、皮膚免疫系の制御と宿主のホメオスタシスへの影響を強調するものである。このような状況における肥満細胞の役割は、微生物との相互作用によって異なることは間違いない。マイクロバイオームの特定の構成要素が肥満細胞の耐性と機能を微調整するメカニズムは、まだ解明され始めたばかりである。宿主と微生物の相互作用という新たな分野は、感染症や炎症性疾患と闘うための新たな治療法の開発に道を開く可能性を秘め、今後の研究に多くのエキサイティングな機会を与えてくれる。
自然免疫細胞の中でも皮膚肥満細胞は、最大12週間という驚くべき長寿を示し、活性化後に顆粒を補充し、変化させる能力を持っている。このユニークな特徴は、病原体や危険信号、潜在的に常在する微生物叢に遭遇したときに、マスト細胞に固有の短期記憶を与える。この記憶概念は、他の自然免疫細胞で観察される「訓練された免疫」に似ており、その後の遭遇時にマスト細胞の反応に影響を与え、防御反応とアレルギー反応の両方に影響を与える可能性がある。この自然免疫記憶を担う根本的なメカニズムには、ヒストン修飾、DNAメチル化、特定のマイクロRNAやノンコーディングRNAの発現といったプロセスを含む、エピジェネティックなリプログラミングが関与していると考えられている。これらのメカニズムは総称して、刺激に応じて細胞の転写プログラ ムを再構築する([95]に総説あり)。しかし、刺激に反応した肥満細胞の機能プログラムの長期的な変化を調べる研究は少ない。
例えば、マスト細胞はLPS刺激後、マクロファージにおけるエンドトキシン耐性と同様の短期記憶を示すことがある。肥満細胞におけるIgEとLPS刺激との相互作用もいくつか観察されている。IgEによって誘導された感作が、NF-κB転写因子の予備活性化を介して、LPSに対するより高い応答に向けて肥満細胞をプライミングする [96]。より最近の研究では、IgEとβグルカンの刺激ではマスト細胞に耐性や訓練が誘導されなかったが、LPS条件付けではシグナル伝達経路に有意かつ持続的な変化が生じたことが明らかにされた。LPSは、PI3K-ACTシグナル伝達経路を障害することによって、二次的なLPS刺激に対して無反応の状態をもたらし、ナイーブなマスト細胞と比較して、NF-κB活性化の減少、TNF-αおよびIL-6放出の減少をもたらした。さらに、LPSで刺激された肥満細胞は、生きたカンジダ・アルビカンスに暴露されると、TNF-α放出の増加を示した。このことは、LPSが、その後のチャレンジに応じて、寛容反応と訓練反応の両方を誘導できることを示唆している。注目すべきことに、LPS刺激中にHDACを阻害すると、二次的なLPSチャレンジに対するLPS刺激を受けた肥満細胞の応答は部分的に回復したが、C. albicansに暴露されたときのサイトカイン産生の増加は回復しなかった [97]。このことは、他の自然免疫細胞と同様に、肥満細胞も自然免疫記憶を発達させることができ、異なる刺激条件が、肥満細胞が局所の炎症反応を減衰させるか増強させるかに影響することを示している。それでもなお、訓練された免疫が様々な刺激に対して肥満細胞に適用されるかどうか、in vivoでの関連性、そしてその基礎となるメカニズムは依然として不明である。特に常在細菌とのクロストークにおいて、訓練された免疫が肥満細胞応答の調節に果たす潜在的役割を理解するためには、さらなる研究が必要である。
著者貢献
構想、N.L.およびP.P.、執筆・原案作成、C.J.B.およびC.G.、執筆・査読・編集、N.L.およびP.P.、視覚化、N.L.およびP.P.、監修、P.P.、資金獲得、P.P.。
資金提供
本研究は、カナダ自然科学・工学研究評議会(NSERC RGPIN-2022-03453)の支援を受けた。
利益相反
著者らは利益相反がないことを宣言する。
参考文献
ホメオスタシスとアトピー性皮膚炎における皮膚微生物叢と上皮-免疫のクロストーク-ミニレビュー。Vet. Dermatol. 2021, 32, 533-e147. [Google Scholar] [CrossRef] [PubMed].
サーモン、J.K.; アームストロング、C.A.; アンセル、J.C. 免疫器官としての皮膚。West. J. Med. 1994, 160, 146-152. [Google Scholar] [PubMed].
皮膚の免疫学的解剖学. Nat. Rev. Immunol. 2019, 19, 19-30. [Google Scholar] [CrossRef] [PubMed].
Voss, M.; Kotrba, J.; Gaffal, E.; Katsoulis-Dimitriou, K.; Dudeck, A. 皮膚における肥満細胞: 皮膚における肥満細胞:完全性の擁護者か、それとも炎症における加害者か?Int. J. Mol. Sci. 2021, 22, 4589. [Google Scholar] [CrossRef] [PubMed].
皮膚微生物叢と免疫の対話。Science 2014, 346, 954-959. [Google Scholar] [CrossRef] [PubMed].
Byrd、A.L.; Belkaid、Y.; Segre、J.A. ヒト皮膚マイクロバイオーム。Nat. Rev. Microbiol. 2018, 16, 143-155. [Google Scholar] [CrossRef].
Dwyer, D.F.; Barrett, N.A.; Austen, K.F. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat. Immunol. 2016, 17, 878-887. [Google Scholar] [CrossRef].
Quaresma, J.A.S. Organization of the skin immune system and compartmentalized immune responses in infectious diseases. Clin. Microbiol. Rev. 2019, 32, e00034-18. [Google Scholar] [CrossRef].
Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289-301. [Google Scholar] [CrossRef].
Zhang, C.; Merana, G.R.; Harris-Tryon, T.; Scharschmidt, T.C. 皮膚免疫: 私たちの体の外側のバリアの複雑な生物学を解剖する。Mucosal Immunol. 2022, 15, 551-561. [Google Scholar] [CrossRef].
Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. 創傷治癒における肥満細胞の寄与に関する総説: 創傷治癒における肥満細胞の貢献に関する総説:関与する分子および細胞メカニズム。Clin. Rev. Allergy Immunol. 2020, 58, 298-312. [Google Scholar] [CrossRef] [PubMed].
抗菌ペプチドによる肥満細胞の活性化におけるGタンパク質共役受容体の役割: 関連性はあるのか?Immunol. Cell Biol. [Google Scholar] [CrossRef] [PubMed].
Pundir, P.; Catalli, A.; Leggiadro, C.; Douglas, S.E.; Kulka, M. 新規抗菌ペプチドであるプレウロシジンはFPRL1受容体を介してヒト肥満細胞の活性化を誘導する。Mucosal Immunol. 2014, 7, 77-87. [Google Scholar] [CrossRef] [PubMed].
Pundir、P.; MacDonald、C.A.; Kulka、M. 新規受容体C5aR2はc5aを介したヒト肥満細胞の接着、遊走、炎症性メディエーター産生に必要である。J. Immunol. 2015, 195, 2774-2787. [Google Scholar] [CrossRef].
Tauber、M.; Basso、L.; Martin、J.; Bostan、L.; Pinto、M.M.; Thierry、G.R.; Houmadi、R.; Serhan、N.; Loste、A.; Bleriot、C.; et al. マウスとヒトにおける臓器間の肥満細胞集団のランドスケープ。J. Exp. Med. 2023, 220, e20230570. [Google Scholar] [CrossRef].
Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajénoff, M. Hemogenic endothelial fate mapping reveals dual developmental origin of mast cells. Immunity 2018, 48, 1160-1171.e5. [Google Scholar] [CrossRef].
Gurish, M.F.; Austen, K.F. 肥満細胞サブセットの発生起源と機能的特殊化。Immunity 2012, 37, 25-33. [Google Scholar] [CrossRef].
Li,Z.;Liu,S.;Xu,J.;Zhang,X.;Han,D.;Liu,J.;Xia,M.;Yi,L.;Shen,Q.;Xu,S.;他。 成人結合組織常在マスト細胞は後期赤血球骨髄系前駆細胞に由来する。Immunity 2018, 49, 640-653.e5. [Google Scholar] [CrossRef].
St John, A.L.; Rathore, A.P.S.; Ginhoux, F. New Perspectives on the origins and heterogeneity of mast cells. Nat. Immunol. 2023, 23, 55-68. [Google Scholar] [CrossRef].
マスト細胞の発生。Proc. Jpn. マスト細胞の発生. Ser. B Phys. Sci. 2007, 83, 164-174. [Google Scholar] [CrossRef].
マスト細胞と鍼治療による鎮痛。Cells 2022, 11, 860. [Google Scholar] [CrossRef] [PubMed].
肥満細胞前駆体の起源、成熟およびリクルートメント。Front. Biosci. (Schol. Ed.) 2011, 3, 1390-1406. [Google Scholar] [PubMed].
神経アレルギー学: 肥満細胞-神経クロストーク. Allergol. 2022, 71, 288-293. [Google Scholar] [CrossRef].
ヒスタミンはin vivoで血流量を増加させ内皮バリアを破壊することにより血管透過性亢進を誘導する。PLoS ONE 2015, 10, e0132367. [Google Scholar] [CrossRef] [PubMed].
炎症性および腫瘍の血管新生およびリンパ管新生における肥満細胞と好塩基球。Eur. J. Pharmacol. 2016, 778, 146-151. [Google Scholar] [CrossRef] [PubMed].
マスト細胞活性化の理解における最近の進歩、あるいはマスト細胞メディエーター障害というべきか?専門家による臨床免疫学(Expert Rev. Clin. Immunol. 2019, 15, 639-656. [Google Scholar] [CrossRef].
Cheng、L.E.;Hartmann、K.;Roers、A.;Krummel、M.F.;Locksley、R.M.血管周囲肥満細胞は、IgEを捕捉するために皮膚血管を動的にプローブする。Immunity 2013, 38, 166-175. [Google Scholar] [CrossRef].
Sumpter, T.L.; Balmert, S.C.; Kaplan, D.H. 樹状細胞と肥満細胞が介在する皮膚免疫応答。JCI Insight 2019, 4, e123947. [Google Scholar] [CrossRef].
幹細胞因子は肥満細胞の活性化表現型をプログラムする。J. Immunol. 2012, 188, 5428-5437. [Google Scholar] [CrossRef].
ラットc-kitリガンドである幹細胞因子による肥満細胞の増殖、成熟、ヘパリン合成の誘導。Proc. Natl. Acad. Sci. USA 1991, 88, 6382-6386. [Google Scholar] [CrossRef].
Artuc, M.; Steckelings, U.M.; Henz, B.M. 肥満細胞と線維芽細胞の相互作用: 線維芽細胞および上皮細胞成長因子の供給源および誘導因子としてのヒト肥満細胞。J. Investig. Dermatol. 2002, 118, 391-395. [Google Scholar] [CrossRef].
PAI1は皮膚線維症における線維芽細胞と肥満細胞の相互作用を媒介する。J. Clin. Investig. 2018, 128, 1807-1819. [Google Scholar] [CrossRef] [PubMed].
ヒスタミンはペリオスチン発現を介して組織リモデリングに寄与する。J. Investig. Dermatol. 2014, 134, 2105-2113. [Google Scholar] [CrossRef] [PubMed].
ヒト黒色腫の自己分泌増殖因子としてのbFGF。Oncogene Res. 1988, 3, 177-186. [Google Scholar] [PubMed].
Schadendorf、D.; Moller、A.; Algermissen、B.; Worm、M.; Sticherling、M.; Czarnetzki、B.M. ヒト悪性黒色腫細胞がin vitroで産生するIl-8は必須の自己分泌増殖因子である。J. Immunol. 1993, 151, 2667-2675. [Google Scholar] [CrossRef] [PubMed].
Hernandez-Barrera、R; Torres-Alvarez、B; Castanedo-Cazares、J.P.; Oros-Ovalle、C; Moncada、B. 肝斑の病因における重要な特徴としての太陽性エラストーシスと肥満細胞の存在。Clin. Exp. Dermatol. 2008, 33, 305-308. [Google Scholar] [CrossRef] [PubMed].
ヒスタミンはin vitroで正常ヒトメラノサイトを刺激する: ヒスタミンはin vitroで正常ヒトメラノサイトを刺激する。J. Dermatol. Sci. 1993, 6, 146-154. [Google Scholar] [CrossRef].
ヒスタミンはヒト正常メラノサイトにおいてH2受容体を介したプロテインキナーゼa活性化によりメラニン生成と形態変化を誘導する。J. Investig. Dermatol. 2000, 114, 334-342. [Google Scholar] [CrossRef].
Wernersson, S.; Pejler, G. 肥満細胞分泌顆粒: 戦いのための武装。Nat. Rev. Immunol. 2014, 14, 478-494. [Google Scholar] [CrossRef].
Komi, D.E.A.; Wöhrl, S.; Bielory, L. Mast cell biology at molecular level: A comprehensive review. Clin. アレルギーの免疫学。2020, 58, 342-365. [Google Scholar] [CrossRef].
McNeil, B.D.; Pundir, P.; Meekar, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature 2015, 519, 237-241. [Google Scholar] [CrossRef] [PubMed].
マウスの構成性結合組織および誘導性粘膜肥満細胞のプロテアーゼ表現型は組織によって制御される。Proc. Natl. Acad. Sci. USA 2011, 108, 14210-14215. [Google Scholar] [CrossRef] [PubMed].
ヒト肥満細胞トランスクリプトームの再定義(Deep-CAGEシークエンシングによる)。Blood 2014, 123, e58-e67. [Google Scholar] [CrossRef] [PubMed].
Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A mast-cell-specific receptor mediates neurogenic inflammation and pain. Neuron 2019, 101, 412-420.e3. [Google Scholar] [CrossRef].
Pundir, P.; Liu, R.; Vasvada, C.; Serhan, N.; Limjunyawong, N.; Yee, R.; Zhan, Y.; Dong, X.; Wu, X.; Zhang, Y.; et al. 結合組織肥満細胞特異的受容体は、細菌のクオラムセンシング分子を検出し、抗菌免疫を媒介する。Cell Host Microbe 2019, 26, 114-122.e8. [Google Scholar] [CrossRef].
モルヒネ髄腔内注入により誘発される肉芽腫には、肥満細胞特異的受容体が重要である。J. Immunol. 2019, 203, 1701-1714. [Google Scholar] [CrossRef].
MRGPRX2は塩酸シノメニンによるアナフィラキシー様反応に必須である。Biochem. Pharmacol. 2017, 146, 214-223. [Google Scholar] [CrossRef].
代表的な抗菌薬は、MRGPRX2およびそのマウスホモログMRGPRB2を介して、肥満細胞の脱顆粒およびアナフィラキシー様反応を誘導する。Eur. J. Immunol. 2017, 47, 1949-1958. [Google Scholar] [CrossRef].
Grice、E.A.; Kong、H.H.; Conlan、S.; Deming、C.B.; Davis、J.; Young、A.C.; NISC Comparative Sequencing Program; Bouffard、G.G.; Blakesley、R.W.; Muray、P.R.; et al. ヒト皮膚マイクロバイオームの局所的および時間的多様性。Science 2009, 324, 1190-1192. [Google Scholar] [CrossRef].
Grice、E.A.; Segre、J.A. 皮膚マイクロバイオーム。Nat. Rev. Microbiol. 2011, 9, 244-253. [Google Scholar] [CrossRef]。
ヒト皮膚マイクロバイオームの時間的安定性。Cell 2016, 165, 854-866. [Google Scholar] [CrossRef] [PubMed].
ヒト皮膚における真菌および細菌群集のトポグラフィー的多様性。Nature 2013, 498, 367-370. [Google Scholar] [CrossRef] [PubMed].
アトピー性皮膚炎の皮膚マイクロバイオーム. Allergol. 2022, 71, 31-39. [Google Scholar] [CrossRef] [PubMed].
Staphylococcus epidermidis espは黄色ブドウ球菌のバイオフィルム形成と宿主-病原体相互作用に関連する特異的タンパク質を分解する。J. Bacteriol. 2013, 195, 1645-1655. [Google Scholar] [CrossRef].
このような研究は、皮膚免疫のコンパートメント制御を常在菌が担っていることを示唆している。Science 2012, 337, 1115-1119. [Google Scholar] [CrossRef].
Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.R.; Hooper, L.V.; Aulock, S.V.; Radek, K.A.; et al. 常在細菌は皮膚傷害後のTLR3依存性炎症を制御する。Nat. Med. 2009, 15, 1377-1382. [Google Scholar] [CrossRef]。
NISC比較シーケンスプログラム(NISC Comparative Sequencing Program); et al. アトピー性皮膚炎患児における疾患再燃と治療に関連した皮膚マイクロバイオームの時間的シフト。Genome Res. 2012, 22, 850-859. [Google Scholar] [CrossRef].
Lewis、D.J.; Chan、W.H.; Hinojosa、T.; Hsu、S.; Feldman、S.R. 乾癬における微生物病原のメカニズムと皮膚マイクロバイオームの役割: 総説。Clin. Dermatol. 2019, 37, 160-166. [Google Scholar] [CrossRef].
Marshall, J.S. Mast-cell responses to pathogens. Nat. Rev. Immunol. 2004, 4, 787-799. [Google Scholar] [CrossRef]。
Yu,Y.、Blockhuis,B.R.、Garssen,J.、Redegeld,F.A. 非IgE介在性肥満細胞活性化。Eur. J. Pharmacol. 2016, 778, 33-43. [Google Scholar] [CrossRef].
マイクロバイオームは正常皮膚の表皮下コンパートメントにまで及んでいる。Nat. Commun. 2013, 4, 1431. [このような研究は、日本ではほとんど行われていない。
Bay、L.; Barnes、C.J.; Fritz、B.G.; Thorsen、J.; Restrup、M.E.M.; Rasmussen、L.; Sorenson、J.K.; Hesselvig、A.B.; Odgaard、A.; Hansen、A.J.; et al. ヒト皮膚における普遍的真皮マイクロバイオーム。MBio 2020, 11, e02945-19. [Google Scholar] [CrossRef] [PubMed].
Wang、Z.; Mascarenhas、N.; Eckmann、L.; Miyamoto、Y.; Sun、X.; Kawakami、T.; Nardo、A.N. 皮膚マイクロバイオームは、ケラチノサイトにおける幹細胞因子(SCF)産生を誘発することにより、肥満細胞の成熟を促進する。J. Allergy Clin. Immunol. 2017, 139, 1205-1216.e6. [Google Scholar] [CrossRef] [PubMed].
國貞 毅; Lu, S.Z.; Yoshida, H.; Nishikawa, S.; Mizoguchi, M.; Tyrrell, L.; Willaims, D.A.; Wang, X.; Longley, B.J. Murine Cutaneous mastocytosis and epidermal melanocytosis induced by keratinocyte expression of transgenic stem cell factor. J. Exp. Med. 1998, 187, 1565-1573. [Google Scholar] [CrossRef].
Huttunen, M.; Aalto, M.L.; Harvima, R.J.; Hormanheimo, M.; Harvima, I.T. 上皮化創および慢性創傷におけるトリプターゼおよびキマーゼ活性を示す肥満細胞の変化: 慢性創傷における肥満細胞とトリプターゼおよびキマーゼ活性。Exp. Dermatol. 2000, 9, 258-265. [Google Scholar].
Canesso、M.C.C.、Vieira、A.T.、Castro、T.B.R.、Schirmer、B.G.A.、Cisalpino、D.、Martins、F.S.、Rachid、M.A.、Nicoli、J.R.、Teixeira、M.M.、Barcelos、L.S. 皮膚創傷治癒は常在細菌叢の非存在下で促進され、瘢痕を残さない。J. Immunol. 2014, 193, 5171-5180. [Google Scholar] [CrossRef].
創傷治癒における肥満細胞の役割。Int. Wound J. 2010, 7, 55-61. [Google Scholar] [CrossRef]。
マスト細胞はマウスの皮膚創傷の正常な治癒に必要である。faseb j. 2006, 20, 2366-2368. [Google Scholar] [CrossRef]。
皮膚微小環境における常在細菌に対する肥満細胞の耐性は線維芽細胞によって制御されている。Cell Rep. 2023, 42, 112453. [Google Scholar] [CrossRef]。
Cayrol、C.; Girard、J.P. インターロイキン-33(IL-33): その生物学と強力な細胞外サイトカインとしての放出に関与するメカニズムの批判的レビュー。サイトカイン2022, 156, 155891. [Google Scholar] [CrossRef].
IL-33はIRAK1分解を誘導することにより、細菌細胞壁産物に対する選択的肥満細胞耐性を引き起こす。Eur. J. Immunol. 2013, 43, 979-988. [Google Scholar] [CrossRef] [PubMed].
非病原性常在性大腸菌は肥満細胞の脱顆粒を抑制する。Exp. Dermatol. 2008, 17, 427-435. [Google Scholar] [CrossRef] [PubMed].
肥満細胞のメディエーター反応と病原性微生物および常在微生物によるその抑制。Mol. Immunol. 2015, 63, 74-79. [Google Scholar] [CrossRef] [PubMed].
肥満細胞を飼いならす微生物: アレルギー性炎症とそれ以外への影響。Eur. J. Pharmacol. 2016, 778, 169-175. [Google Scholar] [CrossRef].
大腸菌と相互作用するマスト細胞は、自然免疫に関連する遺伝子をアップレギュレートし、Fc(ε)RIを介した活性化には反応しなくなる。J. Leukoc. Biol. 2006, 79, 339-350. [Google Scholar] [CrossRef].
マウス骨髄由来肥満細胞の免疫グロブリンEを介した脱顆粒および後期免疫反応に対する加熱死乳酸菌株の抑制効果. Anim. 2010, 81, 714-721. [Google Scholar] [CrossRef].
Forsythe, P.; Wang, B.; Khambati, I.; Kunze, W.A. 摂取した乳酸菌の全身作用: 肥満細胞膜カリウム(IKCa)電流および脱顆粒の抑制。PLoS ONE 2012, 7, e41234. [Google Scholar] [CrossRef].
Oksaharju, A.; Kankainen, M.; Kekkonen, R.A.; Lindstedt, K.A.; Kovanen, P.T.; Korpela, R.; Miettinen, M. プロバイオティクス乳酸菌は、ヒト肥満細胞におけるFCER1とHRH4の発現をダウンレギュレートする。World J. Gastroenterol. 2011, 17, 750-759. [Google Scholar] [CrossRef].
Schiffer, C.; Lalanne, A.I.; Cssard, L.; Mancardi, D.A.; Malbec, O.; Bruhns, P.; Dif, F.; Daeron, M. 乳酸菌カゼイ株は免疫炎症のエフェクター相を抑制する。J. Immunol. 2011, 187, 2646-2655. [Google Scholar] [CrossRef]。
ビフィズス菌はIgEを介したラット好塩基球性白血病(RBL-2H3)細胞の脱顆粒を抑制する。Microbiol. Immunol. 2010, 54, 54-57. [Google Scholar] [CrossRef].
プロバイオティクスLactobacillus johnsonii NCC533(La1)を離乳期の特定の時期に経口投与すると、モデルマウスNC/Ngaにおいて成熟後に誘発されるアトピー性皮膚炎の発症を予防できる。Br. J. Dermatol. 2007, 156, 499-509. [Google Scholar] [CrossRef] [PubMed].
Hacini-Rachinel、F.; Gheit、H.; Le Ludec、J.B.; Dif、F.; Nancey、S.; Kaiserlian、D. 経口プロバイオティクスはエフェクターT細胞と制御性T細胞の両方に作用して皮膚の炎症を制御する。PLoS ONE 2009, 4, e4903. [Google Scholar] [CrossRef] [PubMed].
キムチ由来乳酸菌の経口投与はNC/Ngaマウスのアトピー性皮膚炎を抑制した。J. Appl. Microbiol. 2011, 110, 1195-1202. [Google Scholar] [CrossRef] [PubMed].
ラクトバチルス・ラムノサス(Lactobacillus rhamnosus)GGの摂取はNC/Ngaマウスにおけるアトピー性皮膚炎の発症を抑制する。Clin. Exp. Allergy 2007, 37, 296-303. [Google Scholar] [CrossRef] [PubMed].
Lactobacillus(ロイテリ菌)fn041による制御性t細胞の誘導と腸内細菌叢の調節を介したマウスのアトピー性皮膚炎の予防。Mol. Nutr. Food Res. 2022, 66, e2100699. [Google Scholar] [CrossRef] [PubMed].
また、このような研究は、日本人の皮膚病態の改善にも有効であることが示唆された。Front. Immunol. 2018, 9, 1905. [Google Scholar] [CrossRef] [PubMed].
Kim, W.K.; Jang, Y.J.; Han, D.H.; Jeon, K.; Lee, C.; Han, H.S.; Ko, G. Lactobacillus paracasei KBL382投与は、免疫応答と腸内細菌叢を調節することによりアトピー性皮膚炎を減弱させる。Gut Microbes 2020, 12, 1819156. [Google Scholar] [CrossRef].
de Andrade, P.; E Silva, J.M.; Carregaro, V.; Sacramento, L.A.; Roberti, L.R.; Aragon, D.C.; Carmona, F.; Roxo-Junior, P. アトピー性皮膚炎の小児および青年におけるプロバイオティクスの有効性: 無作為二重盲検プラセボ対照試験。Front. 2021, 8, 833666. [Google Scholar] [CrossRef].
アトピー性皮膚炎の治療と予防のための乳酸菌: 臨床的および実験的エビデンス。Front. Cell. Infect. Microbiol. 2023, 13, 1137275. [Google Scholar] [CrossRef].
Wang、Z.; MacLeod、D.T.; Di Nardo、A. 常在細菌リポテイコ酸は、ワクシニアウイルスに対する皮膚肥満細胞の抗菌活性を増加させる。J. Immunol. 2012, 189, 1551-1558. [Google Scholar] [CrossRef].
Mukherjee, S.; Jemielita, M.; Stergioula, V.; Tikhonov, M.; Bassler, B.L. Photosensing and quorum sensing are integrated to control Pseudomonas aeruginosa collective behaviors. PLoS Biol. [Google Scholar] [CrossRef].
Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. 微生物学(Rev. Microbiol. 2001, 55, 165-199. [Google Scholar] [CrossRef].
細菌のクオラムセンシング: 病原性におけるその役割と制御の可能性。Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef].
カチオン性化合物のMRGPRX2感知-侵害受容と皮膚疾患の橋渡し?Exp. Dermatol. 2021, 30, 193-200. [Google Scholar] [CrossRef].
Monticelli、S.; Leoni、C. 肥満細胞応答のエピジェネティックおよび転写制御。F1000Research 2017, 6, 2064. [Google Scholar] [CrossRef] [PubMed].
IgE依存性感作はLPSに対する応答性を増加させるが、肥満細胞におけるエンドトキシン耐性の発達を修飾しない。Inflamm. Res. 2011, 60, 19-27. [Google Scholar] [CrossRef] [PubMed].
LPSはマスト細胞における訓練と耐性の異なるパターンを誘導する。Front. Immunol. 2022, 13, 835348. [Google Scholar] [CrossRef] [PubMed].
免責事項/出版者注:すべての出版物に含まれる声明、意見およびデータは、著者および投稿者個人のものであり、MDPIおよび/または編集者のものではありません。MDPIおよび/または編集者は、コンテンツで言及されているアイデア、方法、指示、製品に起因する人または財産の損害について、一切の責任を負いません。
著者による© 2023。ライセンシー MDPI, Basel, Switzerland. 本論文は、クリエイティブ・コモンズ 表示(CC BY)ライセンス(https://creativecommons.org/licenses/by/4.0/)の条項および条件の下で配布されるオープンアクセス論文である。
共有と引用
MDPIおよびACSスタイル
皮膚恒常性と免疫におけるマスト細胞-微生物叢クロストークの新たな役割。Cells 2023, 12, 2624. https://doi.org/10.3390/cells12222624
AMAスタイル
皮膚恒常性と免疫におけるマスト細胞-微生物叢クロストークの新たな役割。細胞。2023; 12(22):2624. https://doi.org/10.3390/cells12222624
シカゴ/トゥラビアンスタイル
ボスベルド、キャメロン・ジャクソン、コリン・グース、ナタチット・リムジュニャウォン、プリヤンカ・プンディール。2023. "Emerging Role of the Mast Cell-Microbiota Crosstalk in Cutaneous Homeostasis and Immunity" Cells 12, no. 22: 2624. https://doi.org/10.3390/cells12222624
他のスタイルを探す
出版社名、ジャーナル名、書式名を入力してください。
なお、2016年創刊号より、本誌ではページ番号の代わりに論文番号を使用しています。詳細はこちらをご覧ください。
論文指標
引用
この記事の引用は見つかりませんでしたが、Google Scholarで確認することができます。
論文アクセス統計
記事アクセス統計
記事閲覧数
14. 11月
15. 11月
16. 11月
17. 11月
18. 11月
19. 11月
20. 11月
21. 11月
22. 11月
23. 11月
24. 11月
25. 11月
0
200
400
600
800
ジャーナル統計の詳細については、こちらをクリックしてください。
同一IPアドレスからの複数のリクエストは1ビューとしてカウントされます。
Cells, EISSN 2073-4409, MDPI発行 RSSコンテンツアラート
詳細情報
論文処理料金
請求書を支払う
オープンアクセスポリシー
MDPIへのお問い合わせ
MDPIの求人
ガイドライン
著者の方へ
査読者の方へ
編集者の方へ
ライブラリアンの方へ
出版社の方へ
学会の方へ
学会主催者の方へ
MDPIの取り組み
サイフォーラム
MDPI書籍
Preprints.org
サイリット
サイプロファイルズ
百科事典
JAMS
プロシーディングスシリーズ
MDPIをフォローする
LinkedIn
フェイスブック
ツイッター
MDPIジャーナルからの発行通知やニュースレターを購読する
オプションを選択
メールアドレスを入力してください
購読する
© 1996-2023 MDPI (スイス、バーゼル) 特に断りのない限り免責事項 ご利用規約 プライバシーポリシー
この記事が気に入ったらサポートをしてみませんか?