耐熱性乳酸菌による熱防御プロバイオティクス機能とその因果構造

本文へスキップ記事へスキップ
エルゼビアロゴ
機能性食品ジャーナル
第113巻、2024年2月、106001号
耐熱性乳酸菌による熱防御プロバイオティクス機能とその因果構造
https://www.sciencedirect.com/science/article/pii/S1756464624000033?via%3Dihub

著者リンク オーバーレイパネルを開く伊藤 佳代 a, 宮本 博邦 b c d e, 松浦 真紀子 b c, 石井 千歳 c e, 中西 由美子 e, 須田 渉 e, 佐藤 隆 f, 本田 冬子 a, 黒谷 淳 g、 辻直子 c、中熊輝乃 b c d、進藤千恵 e、伊藤俊幸 h、道端亮之介 i、山田亮一 i、高橋洋介 i、児玉裕明 b c、菊池純 j、大野裕史 e
もっと見る
概要
シェア
引用
https://doi.org/10.1016/j.jff.2024.106001
権利とコンテンツの取得
クリエイティブ・コモンズ・ライセンス
オープンアクセス
ハイライト
プロバイオティクスであるWeizmannia coagulansはブロイラーの熱防御成長を改善する。
腸内糞便と肝臓のオミックスデータから主成分を選択した。
SEMとDirectLiNGAMにより、MLで選択された成分に基づく最適なグループを推定。
熱防御増殖はピリドキサール-オキサレートネットワークと計算上リンクしている。
糞便性シュードモナスは熱防御増殖に負の因果関係を持つ。
要旨
地球温暖化は喫緊の課題であり、人間生活や畜産を管理するために熱ストレス保護物質の開発が必要である。ここでは、耐熱性プロバイオティクス乳酸菌であるWeizmannia coagulans SANK70258の熱防御効果について、人工温熱気候施設におけるブロイラーモデルを用いて検討した。プロバイオティクスへの曝露は、熱ストレス下でもその成長を著しく促進する。機械学習(ランダムフォレストとXGBoost)を用いて選択された特徴的なオミックスデータに基づき、熱ストレス下でのブロイラーの成長に関する構造方程式と因果推論を行った結果、寒冷感受性の原因となる肝シュウ酸の還元に関与するピリドキサールとの強い関係が推定された。また、ピリドキサールを減少させる糞便性シュードモナスは、プロバイオティクスへの暴露によって減少し、成長に負の因果関係を持つと推定される。従って、これらの観察結果は、寒冷感受性に関連する耐熱性のメカニズムを調節するW. coagulansの新しいプロバイオティクスの側面を示唆している。
グラフィカル抄録
ダウンロード 高解像度画像ダウンロード(215KB)
ダウンロード フルサイズ画像のダウンロード
前の記事次の記事
キーワード
熱ストレスプロバイオティクス成長ブロイラー
はじめに
気候変動に関する政府間パネル(IPCC)によると、地球温暖化の問題は極めて深刻な問題であり、生態系全体を脅かし(Stocker et al. 高温条件下で健康を維持し、恒常性を維持するために、生物は外気温の上昇に耐えるメカニズムを進化させてきたが、生体にかかる負担は甚大である(Cramerら、2022)。熱ストレスは畜産業で問題となっており、多くの熱波が死亡率の増加による壊滅的な経済損失をもたらしている。熱ストレスはまた、抗生物質の使用を必要とする感染症を引き起こすこともある(Gathiramら、1987年)。抗生物質の過剰使用は世界的な問題であり、家畜における抗生物質の過剰使用や誤用による抗生物質耐性菌の出現は世界的に明らかになっている。世界保健機関(WHO)は抗菌薬耐性(AMR)に対する行動計画(Holloway et al. また、最近の因果関係研究では、抗生物質の使用は免疫系の機能や温室効果ガスであるメタンの発生に悪影響を及ぼすと推定されている(Okada et al.) これらの問題、すなわち地球温暖化対策と抗生物質の使用削減は、それぞれ別の研究分野であるように見えるが、どちらも人間と動物の食物連鎖と健康連鎖に悪影響を及ぼす。
したがって、熱ストレスが動物に及ぼす影響とその対策に関する研究は不可欠である。比較的管理しやすい家畜を使って、行動や生理的影響を評価する研究が盛んに行われている。牛(West, 2003)、豚(Liuら, 2022)、鶏(Brugalettaら, 2022)の生産に対する熱ストレスの悪影響は、以下のように調査されている:行動変化(例:無気力、飼料摂取量の減少、パンティング)、代謝(例:異化状態、脂肪蓄積、脂肪蓄積、脂肪蓄積、異化状態、脂肪蓄積、異化状態)、代謝(例:異化状態、異化状態、脂肪蓄積、異化状態)、代謝(例:異化状態、異化状態、脂肪蓄積、異化状態)、 代謝(異化状態、脂肪蓄積、骨格筋付着力の低下など)、ホメオスタシス全般(アルカローシス、ホルモンバランスの乱れ、免疫不全、炎症、酸化ストレスなど)、消化管機能(消化器系および吸収障害、腸炎、副細胞バリア障害、嚥下障害など)。そのため、暑熱ストレスに対する生理学的指標を把握するために、以下のような様々なアプローチが研究されている:牛の機械学習を用いた暑熱ストレスの重症度を評価するスコアリングシステムの開発(Beckerら、2021年);牛の暑熱ストレスの影響を軽減するための子牛冷却の必要性(Laporta、2021年);暑熱ストレス時の牛の免疫指標(Bagathら、2019年);牛の繁殖成績の指標(Dauriaら、 2022年);精液を用いた豚の熱ストレス影響評価法の開発(Heら、2020年、Ishidaら、2022年);鶏の脳反応における遺伝子発現パターン(Brugalettaら、2022年、Goelら、 2021);ニワトリの慢性熱ストレスに対する小腸内の炎症性サイトカインおよび抗炎症性サイトカインのレベル(Siddiquiら、2020);寄生虫感染を伴う場合の指標(Schneidersら、2020)。これらの結果からわかるように、遺伝子発現への影響については、遺伝的要因だけでなくエピジェネティックな要因も含めて多くの研究が進んでいる。
一方、これらの課題を克服するために、家畜における栄養素投与の効果に関する研究が進められている。例えば、抗酸化活性を有するビタミンCの投与(Pardue et al., 1985)、ジヒドロミリセチン フラボノイドの投与はミトコンドリア機能障害を軽減し、熱ストレスによる上皮細胞のアポトーシスを抑制する(Wang et al、 2021)、抗酸化物質の生成を助けるセレン(Khan et al., 2019)やα-リポ酸(Wasti et al., 2021)を栄養補助食品として投与することで、家畜の死亡率や熱ストレスによる悪影響を軽減することができる。家畜におけるプレバイオティクスやプロバイオティクス投与の効果に関する研究も進んでいる。プロバイオティクスやプレバイオティクスは、熱ストレスによって引き起こされる腸や肝臓の循環の悪影響を改善する傾向があり(Ringseis and Eder, 2022, Salem et al., 2022)、枯草菌の単独使用(T. A. Cramer et al., 2018)や乳酸菌との併用(Song et al., 2014)に関する研究では、ブロイラーの腸のバリア機能を改善する可能性が報告されている。しかし、地球温暖化が進む中、このような研究が必ずしもブレークスルーにつながるとは限らず、これまで開発されてきた有用素材との併用も含め、新たな技術や視点が求められている。
ここでは、人工的に制御された温熱環境条件下で飼育されたブロイラーモデルを用いて、芽胞形成性プロバイオティクス投与が温熱環境適応性に及ぼす影響を推定した。近年、有胞子性プロバイオティクスの医療への応用が期待されている(Wautersら、2021年)。なかでも、1949年(中山、1950年)から伝統的に食品や飼料に利用されてきた、わずかな環境変異を持つ熱安定性の高い有胞子性乳酸菌であるWeizmannia coagulans(シノニム:Bacillus coagulans)SANK70258をプロバイオティクスとして利用した。通常の条件下で得られたこれまでの知見(Ara et al., 2009, Ito et al., 2022, Sasaki et al., 2020, Watanabe et al., 2018)から、腸内の代謝機能への影響が示唆されたため、腸内糞便と肝臓のオミックス解析を計算機的に行った。その結果に基づいて、2種類の機械学習(ML)法を適用し、オミックスデータから耐熱性に関連する潜在的に重要な特徴成分を抽出した。これらのネットワーク構造の特徴は、共分散構造解析と因果推論によって統計的に評価された。その結果、W. coagulans投与によるいくつかの効果について既知の傾向が検証された。また、ピリドキサール-シュウ酸(シュウ酸)代謝ネットワークが、耐寒性に関連する因子群の中で役割を果たしている可能性が新たに観察された。これらの推定は、耐熱性の分子メカニズムの一端を示すものであり、今後、環境ストレスと消化器病学との関係の解明に役立つものと思われる。
結果
2.1. 本研究のスキームと育種結果
本研究の給餌試験の概要と解析の流れを図 1 に示す。図 1a および S1 に示すように、ブロイラーは 25 °C から 32 °C の人工的に制御された高温で飼育された。図1bに示すように、10日目までは1ケージで飼育し、10日目以降は1ケージに3羽ずつ飼育した。21日後、1ケージあたり1羽を繁殖させた。人工的な温度管理のため、飼育管理はヒヨコ専用ハウス(バッテリーブルーダー)で10日間行った。この条件下での繁殖結果を表1に示す。
ダウンロード 高解像度画像ダウンロード(507KB)
ダウンロード フルサイズ画像のダウンロード
図1. 本研究の概念図。(a)本研究の目的は、通常気象条件下および人工的な暑熱ストレス飼育条件下における耐熱性プロバイオティクスの効果を検討することである。ブロイラーは人工気象施設(ズートロン)で暑熱ストレス条件下で飼育した。詳細な暑熱管理スケジュールをFig.S1に示す。特に、プロバイオティクスW. coagulans(Weizmannia coagulans SANK70258)の効果を評価した。(b)実験計画を以下に示す:0日目から10日目まではバタリーハウスで飼育し、10日目から21日目まではケージで3羽飼育した。その後、大きくなった21日目からは、1羽につき1つのケージで飼育した。49日目の剖検で得られたサンプルをオミックス解析に用いた。(c)本研究の解析手順を示す。飼育成績を確認後、大腸糞便と肝臓のオミックス解析を行った。これらのデータをもとに、構造方程式モデリングと因果推論を行い、飼育成績との因果構造を評価した。
表1. ブロイラー鶏の成長データ。ADGは1日平均体重増加率、F/Gは表示された制限期間中の成長に対する飼料効率を示す(*、p < 0.05)。
項目 空房 常温 高温
空房 N-コントロール H-コントロール H-コアギュランス
バッテリーブルーダー
体重 (n = 12-13) Day0 44.13 ± 1.01 (12) 44.15 ± 0.76 (13)
Day10 243.96 ± 11.88 (12) 271.31 ± 12.15 (13)
アドバンテージ 19.98 ± 1.18 (12) 22.72 ± 1.13 (13)
f/g 0.85 ± 0.06 (12) 0.93 ± 0.06 (13)
通常および熱ストレス条件
体重 (n = 6)(n = 3/ケージ) Day10 246.42 ± 15.72 241.50 ± 19.29 277.67 ± 14.39
21日目 920.00 ± 45.17 921.67 ± 63.11 1003.33 ± 38.70
61.23 ± 2.93 61.83 ± 4.05 65.97 ± 2.31
f/g 1.28 ± 0.06 1.22 ± 0.09 1.31 ± 0.05
体重(n = 5)(n = 1/ケージ) Day21 920.00 ± 45.17 921.67 ± 63.11 1003.33 ± 38.70
Day49 3998 ± 216.83 3756.0 ± 94.11 4117.0 ± 61.27
ADG 105.28 ± 6.78 95.86 ± 3.86 106.24 ± 2.28
f/g 1.66 ± 0.02 1.75 ± 0.06 1.68 ± 0.03
同一個体(n = 5)のまとめ
ADG 10-49日目 93.69 ± 5.29 87.46 ± 2.46 95.71 ± 1.53 * f/g 1.58 ± 0.02 1.75 ± 0.06 1.68 ± 0.03
f/g 1.58 ± 0.01 1.61 ± 0.04 1.59 ± 0.02
合計
ADG Day0-49 80.65 ± 4.44 (5) 75.76 ± 1.94 (5) 83.08 ± 1.25 * (5)
f/g 1.54 ± 0.01 (5) 1.56 ± 0.03 (5) 1.55 ± 0.02 (5)
その結果、10日目に飼育を中止したが、各群間に有意差は認められなかった。加熱処理10日後のデータによると、21日目と比較して有意差はないものの、加熱ストレス試験群(H_coagulans)は加熱ストレス対照群(H_control)に比べて体重が増加する傾向にあった。 21日目から49日目にかけて、ヒートストレス試験群(H_coagulans)はヒートストレス対照群(H_control)に比べて体重が有意に増加した(p < 0.05)。飼料効率も改善する傾向にあった。同様の傾向は、非加熱ストレス条件下における従来の対照群(N_control)でも観察された。これは、10日目から49日目までの加熱処理下と、0日目から49日目までのすべての飼育期間中の両方で確認された。
図1cに示すように、摂食成績が評価され、49日目の剖検時に得られたサンプルについて、大腸内容物および肝臓組織のオミックス解析が行われた。総合的なデータはMLによって分類された。さらに、構造方程式モデルを用いて作用機序を予測し、飼料摂取量に関連する統計的因果構造を因果推論した。
2.2. 腸内容物および肝臓のメタボローム解析
糞便メタボローム解析と肝臓メタボローム解析の結果から、p値0.05以下(高温条件下での2群比較による)の物質をヒートマップ上に示した(図2)。糞便では、3つの条件下で強い変化の傾向が観察され、例えば、熱ストレス下ではより大きな物質の減少が観察され、肝臓でも同様の結果が観察された。p値が0.01以下の物質のうち、糞便ではガラクトース、ラクチトール、シチジン、セバシン酸が増加し、3-ヒドロキシプロピオン酸、アデニンが減少し、肝臓ではエリスルロース、2-アミノイソ酪酸、グルコース-6-リン酸が減少した。肝内循環は生体の恒常性維持に重要である。相関分析は、腸内代謝物と肝臓代謝物のバランスを評価するために行われる。これらの代謝物の相関を評価することで、腸肝循環の傾向を推測した。その結果、腸内容物と肝臓代謝物の相関は、3つの条件下で明らかに異なっていた。その結果、正常群(N_control)(Fig. S2)、熱ストレス対照群(H_control)(Fig. S3a)、熱ストレス試験群(H_coagulans)(Fig. S3b)の間で大きな差が見られた。特に、大腸内の糞便中糖質と正の相関を示す代謝物の濃度は、加熱処理下の熱ストレス対照群(H_control)(図S3a)では低く、正常群(N_control)では増加した(図S2)。熱ストレス試験群(H_coagulans)では、熱ストレス条件下でさらにレベルが上昇した(図S3b)。代謝機能を予測するため、さらにパスウェイ解析を行った(表S1およびS2)。熱ストレス試験群(H_coagulans)の特徴を理解するため、統計的に有意(p < 0.05)で、低濃度(≥0.1 μmol/g)ではない代謝物について解析を行った。図S4に示すように、通常群ではコントロール(N_control)、熱ストレス試験群(H_coagulans)ではガラクトース代謝、アミノ糖・ヌクレオチド糖代謝、デンプン・スクロース代謝、トリプトファン代謝が活性化する傾向が見られた。
ダウンロード 高解像度画像ダウンロード(445KB)
ダウンロード フルサイズ画像のダウンロード
図2. 大腸および肝臓における糞便のメタボロミクス。(a)N_control群(温度管理なしの通常群)(灰色)、H_control群(ズートロンで管理した群)(青色)、H_coagulans群(ズートロンでW.coagulansを投与した群)(緑色)における糞便中のアミノ酸、炭水化物、およびその他の代謝物の代謝プロファイル、(b)各群における肝臓中のアミノ酸、炭水化物、およびその他の代謝物の代謝プロファイル。各クラスターにおける代謝物の相対量を示すヒートマップ(p < 0.2)。アスタリスクはp < 0.01を示す。略号は以下の通り: LF_は大腸の糞便、LV_は肝臓。(この図の凡例中の色に関する言及の解釈については、読者はこの論文のウェブ版を参照されたい)。
これらの結果から、熱ストレス条件下における熱ストレス対照群(H_control)の代謝系は、通常条件下における通常群(N_control)および熱ストレス条件下における試験群(H_coagulans)の代謝系と比較的類似していることが示唆された。試験群(H_coagulans)は、ビタミンB6代謝とフルクトースおよびマンノース代謝に特徴があった。
2.3. 主要代謝物の抽出とネットワークモデルの開発
これらのメタボロームデータをもとに、MLを用いて必須因子を抽出した。その結果を図3に示す。ランダムフォレスト解析の結果、大腸排泄物(図3a)と肝臓(図3b)において、特徴量の指標となるMeanDecreaseGini値が分類された。大腸の糞便では、炭水化物の順位が高かった。肝臓では、解糖経路に関与するグルコース-6-ホスファターゼなどの有機酸が上位にランクされた。XGBoost分析では、特徴量の指標となる重要度は、糞便と肝臓のピリドキシン、マンニトール、シュウ酸のみが高かった(図3c)。
ダウンロード 高解像度画像ダウンロード(516KB)
ダウンロード フルサイズ画像のダウンロード
図3. MLによる特徴因子の抽出。ランダムフォレストにより(a)糞便代謝物、(b)肝臓代謝物の特徴因子(MeanDecreasGini値)を抽出。(c)XGBoostにより選択された特徴因子(重要度)を可視化したもの。略号は以下の通り: LF_は大腸糞便、LV_は肝臓。
3群のランダムフォレストから抽出した代謝物濃度のみを比較した結果(図S5)、糞便中の炭水化物としてガラクトースとラクチトール、有機酸としてアセチルグリシン、核酸の構成成分として3-ヒドロキシプロピオン酸、3-ヒドロキシイソ酪酸、アデニンの挙動は、平常時(N_対照)と熱ストレス試験群(H_コアギュランス)で類似していた。有意差はあったが、ガラクトースとラクチトールは両群で増加する傾向があり、アセチルグリシン、3-ヒドロキシプロピオン酸、3-ヒドロキシイソ酪酸、アデニンは減少する傾向があった。セバシン酸は、糖質、マルチトール、有機酸のうち、熱ストレス試験群(H_coagulans)でのみ有意に増加した。また、肝臓中の代謝物レベルは、正常群(N_対照)と熱ストレス群との間で必ずしも類似していなかった(図S6)。試験群(H_coagulans)では、2-アミノ酪酸とエリスルロースが有意に減少した。グルコース-6-リン酸レベルの変化は、通常状態(N_control)と熱ストレス試験群(H_coagulans)で有意に減少した。
ランダムフォレスト解析とXGBoost解析の両方で同定された特徴的な因子は、炭水化物として糞便マンニトール(p = 0.0007)、ビタミンB6として糞便ピリドキサール、有機酸として肝シュウ酸(p = 0.001)であった(図4a)。3群間の濃度を比較すると、マンニトールとピリドキサルは熱ストレス試験群(H_coagulans)の糞便でのみ有意に増加していた。
ダウンロード 高解像度画像ダウンロード(453KB)
ダウンロード フルサイズ画像のダウンロード
図4. ML、SEM、および因果推論によって検出された必須代謝因子。(a) MLにより選択された代謝物の検出度。(b)選択された代謝物のSEM結果を可視化した。最適なSEM結果は標準化β係数で報告されている。略号は以下の通り: Man:糞便マンニトール;Pyr:糞便ピリドキサール;Oxa:肝臓シュウ酸;W.coa:W. coagulans; Chisq, カイ二乗χ2; p value, p値(カイ二乗); CFI, 比較適合指数; TLI, Tucker-Lewis 指数; RMSEA, 二乗平均平方根近似誤差; SRMR, 標準化残差平方根; GFI, 適合度指数; AGFI, 調整済み適合度指数。緑と赤はそれぞれプラスとマイナスの効果を示す。選択された適合指標はパス内および表S3に示されている。(c) 有向無サイクルグラフ(DAG)は、DirectLiNGAMによって環境条件とともに計算された。DAG 内の各数値は、算出された因果値を示す。略号は以下の通り: LF_は大腸の糞便、LV_は肝臓。(この図の凡例における色の言及の解釈については、読者はこの論文のウェブ版を参照されたい)。
これらの分類に基づき、構造方程式モデリングと因果推論は、これらのグループ間の明確な因果関係を確立するためではなく、因子グループ間の関連性の程度を評価するために用いられた。さらに、統計的手法を選択する際には、分布がガウス分布であるかどうか、等分散が得られているかどうかを評価した(図S7)。3つの因子の計算では、ガウス分布または非ガウス分布のデータが混在していた。そこで、ガウス分布または非ガウス分布を仮定した統計モデルで評価を行った。
ガウス分布を仮定した最尤法の最適値を示す構造式を計算した結果(表S3および図4b)、W. coagulans投与は糞便マンニトールおよび増殖と正の関係を示し、W. coagulans投与は肝シュウ酸と負の関係を示した。糞便中のピリドキサルはW.coagulansと正の関係にあった。
次に、非ガウス分布を仮定し、独立因果分析を適用して環境因子との関係を評価した。その結果(図4c)、熱ストレスと生育が正の関係にある代謝産物はマンニトールとピリドキサールであり、熱ストレスと生育が負の関係にある代謝産物はシュウ酸であった。
これらの結果は、個々の代謝産物の有意な増減ではなく、糞便中のマンニトールとピリドキサール、およびグループとしての肝シュウ酸の関係が、W. coagulansの投与によって影響を受けることを計算上示唆していた。
2.4. 糞便細菌群への影響
次に、大腸の糞便中の細菌集団を分析した。細菌叢の多様性(図S8aおよびS8b)および門レベル(図S8c)では、統計的に有意な差はなかった。その結果、植物叢の多様性(図S8aおよびS8b)および門レベル(図S8c)では、統計的に有意な差は見られなかった。属レベルでは、加熱処理下で乳酸菌の減少傾向が観察された(図5a)。
ダウンロード 高解像度画像ダウンロード(577KB)
ダウンロード フルサイズ画像のダウンロード
図5. 細菌集団とそのSEM、および原因推定。(a)本研究における細菌属の相対存在量。(b)本研究で変化する傾向があった特徴的な細菌(p < 0.2)。(c) 選択した細菌を用いたSEM結果を可視化した。最適なSEM結果は標準化β係数で報告されている。略号は以下の通り: Flv, Flavonifractor; Lct, Lactobacillus; Psd, Pseudomonas; Rmb, Romboutsia; W. coa, W. coagulans; Chisq. coagulans; Chisq, カイ二乗χ2; p value, p値(カイ二乗); CFI, 比較適合指数; TLI, Tucker-Lewis 指数; RMSEA, 二乗平均平方根近似誤差; SRMR, 標準化残差平方根; GFI, 適合度指数; AGFI, 調整済み適合度指数。緑と赤はそれぞれプラスとマイナスの効果を示す。選択された適合指標は、パス内および表S4に示されている。(d)DAGはDirectLiNGAMによって環境条件とともに計算された。DAGの各数値は計算された因果値を示す。(この図の凡例における色の言及の解釈については、読者はこの論文のウェブ版を参照されたい)。
Pseudomonas属の減少傾向、Flavonifractor属とRomboutsia属の増加傾向(図5bとS9a)が、特に熱ストレス試験群(H_coagulans)で観察されたが、p値が0.2以下の有意な変化を示したサンプルは少なかった。4因子の相関を評価したところ、Lactobacillus属とFlavonifractor属の間に負の相関が観察された(図S9b)。また、データはガウス分布と非ガウス分布が混在していたため(図S10)、ガウス分布または非ガウス分布を仮定した統計モデルで評価を行った。まず、ガウス分布を仮定した最尤法で構造式を計算した(表S4)。最適値を示したモデルでは、W.coagulansの投与はRomboutsia属およびFlavonifractor属と正の関係を示した。W.coagulansの投与はPseudomonas属と負の関係を示し、成長と負の関係を示した(図5c)。次に、非ガウス分布を仮定し、環境因子との関係を独立解析により計算評価した(図5d)。その結果、Lactobacillus属とFlavonifractor属は成長と強い関係を示さなかった。Romboustia属では正の成長値(値=6.53)が観察されたが、Pseudomonas属ではより負の成長値(値=-24.99)が観察された。
これらの結果は、W. coagulansの投与により、個々の細菌群の増減ではなく、グループとしての細菌群の関係が変化することを計算上示唆していた。
2.5. オミックスデータの俯瞰図
本研究の3群のうち、熱ストレス試験群(H_coagulans)の飼育成績は、熱ストレス対照群(H_control)よりも有意に良好であり(p < 0.05)(表1)、常温条件下では通常群(N_control)と同等の成績であった。MLおよび統計的構造モデル評価により、3群間の作用機序の大きな違いが推定された(図6)。データを俯瞰してみると(図6a)、代謝物レベルでは、大腸糞便中のマンニトールは、暑熱ストレス対照群(H_対照)を除いて共通であったが、大腸糞便中のマンニトールレベルには若干の差があった。大腸糞便中のピリドキサルは熱ストレス条件(H_対照群およびH_コアギュラン ス群)に馴染んでおり(図6a)、肝臓中のシュウ酸は熱ストレス試験群(H_コアギュラン ス群)のみ影響が少ない傾向にあった(図6a)。細菌叢レベルではRomboustia属の存在量に差があったが(図6b)、それは熱ストレス試験群(H_coagulans)と熱ストレス条件下での対照群(H_control)で観察された。これらの実験条件下では、熱ストレス試験群(H_coagulans)では、熱ストレス対照群(H_control)に比べて、シュードモナス属の影響が極めて小さい傾向にあった(図6b)。したがって、これらの観察結果は、糞便代謝物および微生物叢レベルにおけるプロバイオティクス投与の効果が、熱ストレス条件下で統計学的に観察されたことを示唆している。
ダウンロード 高解像度画像ダウンロード(354KB)
ダウンロード フルサイズ画像のダウンロード
図6. 図4、図5に示したSEM推定成分の寄与度を(a)代謝物、(b)腸内細菌で可視化した。これらから得られたオミックスデータを、各中央値に基づいて二値化することで以下のように整理した: H_controlは常温対照群、H_controlは高温条件下群、H_coagulansは高温条件下でのW. 各代謝物の寄与が異なるため、寄与円の矢印の色は3群で異なるように調整した。略号は以下の通り: LF_は大腸糞便、LV_は肝臓。
考察
本研究は、人工的に制御された熱環境条件下におけるW. coagulansの有胞子性プロバイオティクスとしての熱防御機能の一面を明らかにすることができた。作用機序を推定するため、腸内糞便と肝臓のオミックスデータを、ランダムフォレストとXGBoostの2種類のML手法を用いて分類した。さらに、構造方程式モデリングと因果推論を用いて、最適値を示す因子群をグループとして予測した。複数の環境条件下におけるオミックスデータから代謝機能を推定する試みが必要である可能性が示唆されているが、機械学習アルゴリズム、共分散構造解析、および因果推論を用いて、家畜モデルに対する耐熱効果の作用機序を推定したのは、我々の知る限りこれが初めての試みである。通常条件下で知られているW. coagulans投与による成長促進機能傾向(Itoら、2022)は、暑熱ストレス条件下でも観察された。重要な点は、ピリドキサール-シュウ酸(シュウ酸)代謝ネットワークが、冷感抵抗性に関連する因子群の一翼を担っている可能性があることである。
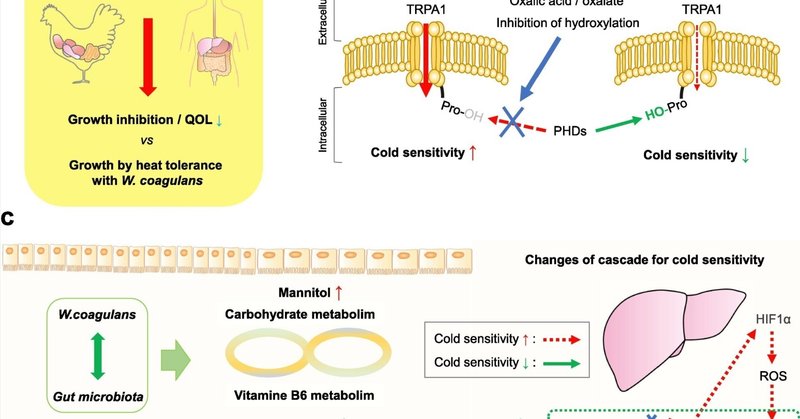
本研究の結果から、W. coagulansの経口投与は耐熱性を制御する可能性があるが、熱ストレスは上記のように動物の成長阻害とQOLに悪影響を及ぼす(図7a)。これらのメカニズムは明言できない。しかし、シュウ酸が、一過性受容体電位アンキリン1(TRPA1)によるプロリン水酸化を阻害することにより、活性酸素種(ROS)と温度に対する感受性を高めることは特筆すべきことである(Miyake et al、 Kurganovら, 2017, Miyakeら, 2016)(図7b)。TRPA1は、動物種を超えていくつかの感覚プロセスに関与しており(Kurganov, Saito, Tanaka Saito, & Tominaga, 2017)、これまでに報告されているように、味覚や温度の感知もそのひとつである。シュウ酸の蓄積は生体に害を及ぼすことが知られている。尿路結石の原因となるシュウ酸はグリオキザール酸から生成される。シュウ酸濃度が高いのは原発性高シュウ酸尿症1型(PH1)で、不溶性のシュウ酸カルシウムが蓄積して腎不全が進行するのが特徴である。PH1の原因は、ピリドキサール5′-リン酸(PLP)依存性酵素であるグリオキシル酸アミノトランスフェラーゼ(AGT)の欠損である(Cellini et al.) このシュウ酸産生を制御する方法として、ピリドキサールまたはピリドキシンによる代謝カスケードの結果、シュウ酸が増加することなくグリシンが形成されることが一般的に知られている (Varalakshmi & Richardson, 1983)。本研究では、MLおよび統計的構造方程式モデル(図4)から、大腸の糞便中ピリドキサールを重要因子の1つとして選択した。ピリドキサールに類似した糞便中ピリドキシンは濃度が低いためML解析から除外されたが、その統計値は熱ストレス試験群(H_coagulans)で有意に高かった(p = 0.0262)(図S11)。したがって、熱ストレス試験群(H_coagulans)の肝臓中のシュウ酸濃度が低いことは、糞便中に検出された高レベルのピリドキサールおよびピリドキシンと関連していると考えるのが妥当である。さらに、熱ストレス試験群(H_coagulans)の腸におけるシュードモナス属の存在量が少ないことは、糞便中のピリドキサールおよびピリドキシンを濃縮する要因の候補として期待される。シュードモナス属はピリドキサールキナーゼを持つことが知られている。この反応では、ATPとピリドキサルが反応し、ADP、H2、ピリドキサール5′-リン酸(https://www.uniprot.org/uniprotkb/J2RGI2/entry)を形成する(Brown et al., 2012)。つまり、腸内にシュードモナスなどのピリドキサールキナーゼを持つ細菌が存在する場合、熱ストレス条件下でW. coagulansによるこのような耐熱効果が生じる可能性がある。
ダウンロード 高解像度画像ダウンロード(638KB)
ダウンロード フルサイズ画像のダウンロード
図7. 本研究から推定される概念図。パネル(a)は、暑さで弱ったニワトリとヒト、すなわち成長阻害と生活の質(QOL)を示している。(b)は、既報(Miyake et al., 2016)の通り、シュウ酸による温度感受性センサーであるtransient receptor potential ankyrin1(TRPA1)の水酸化制御を示している。シュウ酸が存在すると、TRPA1のプロリンリン酸化が阻害され、その結果、温度に対する感受性が亢進することがすでに示されている。この反応には、プロリルヒドロキシラーゼ(PHD)が関与している。赤矢印はイオン流入、緑は水酸化反応、青線は阻害作用を示す。破線はそれぞれの強度が実線よりも弱いことを示す。(c)熱ストレス条件下でのW. coagulansの作用メカニズムを示す推定モデルを可視化した。W. coagulansによる処理後、腸内細菌叢の相互作用に基づき、糖質代謝におけるマンニトールのレベルが上昇した。このモデルでは、ビタミンB6の代謝においてシュードモナスが減少し、ピリドキサルが増加した。この関係の効果は、肝臓におけるシュウ酸の減少であることが示された。さらに、シュウ酸の減少は、PHD(HIF-1α特異的プロリルヒドロキシラーゼ)、低酸素下でのHIF(低酸素誘導因子)1α、酸化ストレスとしての活性酸素種(ROS)、一過性受容体電位アンキリン1(TRPA1)に関係している可能性がある。これらの条件は、以前に報告されたように(Miyake et al., 2016)、寒さに対する感受性の変化を引き起こす過敏性を制御している。赤矢印は、寒冷感受性の亢進のカスケードを示す。破線は、それぞれの強度が実線よりも弱いことを意味し、本研究の熱ストレス条件下でのW. coagulansの曝露によって引き起こされた弱いカスケードを示す。また、緑色の矢印は、本研究の熱ストレス条件下におけるW. coagulansの作用機序として、寒さに対する感受性の低下という推定されるカスケードを示している。(この図の凡例における色に関する言及の解釈については、この論文のウェブ版を参照されたい)。
この文脈において、ピリドキシン-シュウ酸の代謝に関与する代謝物の挙動を、必ずしも有意差のない低濃度(<0.1μmol/g)で存在する代謝物を含めて推定した(図S11)。ATPとピリドキサルは、AGTの酵素反応によってADP、水素、PLPに変換される。今回のメタボローム解析では、PLP濃度は検出下限値以下であった。アデノシン三リン酸(ATP)とアデノシン二リン酸(ADP)は測定できなかったが、糞便アデニン濃度は熱ストレス試験群(H_coagulans)で低下する傾向があった(p = 0.0588)。グリオキシル酸は前述のようにピリドキサールによってグリシンに変換されることから(Varalakshmi & Richardson, 1983)、糞便中および肝臓中のグリオキシル酸およびグリシン濃度を確認した(図S11)。その結果、熱ストレス試験群(H_coagulans)では、肝臓のグリオキシル酸濃度は少なくとも肝臓では減少する傾向が見られたが(p = 0.1185)、グリシンの濃度は有意な変化は見られなかった。遺伝子組換え動物に関する研究は行われていないが、ブロイラー鶏モデルにおいて、糞便中のビタミンB6(ピリドキシンおよびピリドキサール)の増加と肝臓のシュウ酸の減少が有意であったことは重要な現象と考えられる。このように、熱ストレス試験群(H_coagulans)では、過剰な酸化ストレスを緩和する反応のためのシュウ酸の含量が減少していることが注目された。
以上の厳選されたオミックスデータの組み合わせから、本研究から導かれた仮説が図7に可視化された。熱ストレスは生物の成長、免疫、生殖活動、QOLを阻害するが(Bagath et al., 2019, Schneiders et al., 2020, Siddiqui et al., 2020)、W. coagulansを併用すると熱ストレスに強くなった(図7a)。この研究で予測されたメカニズムは、図7bと7cの概念図に示されている。すなわち、W. coagulans投与後の増殖とマンニトールの増加は、既報(Ito et al., 2022)と同様に、効果的な糖質利用の指標であると想定される。第二に、シュードモナス属の還元現象が、上述のようにそのPLPの酵素反応を減衰させ、その結果、腸内でのピリドキサール/ピリドキシンの還元を防ぐと予想される(https://www.uniprot.org/uniprotkb/J2RGI2/entry)。実際、W. coagulansの投与によってシュードモナス属が減少すると、腸内のピリドキサール/ピリドキシン濃度が高くなった(図4aおよびS11)。これらの反応を踏まえると、上述のようにシュウ酸はTRPA1の反応を負に制御することから(Miyake et al. この仮説を支持するため、プロリルヒドロキシラーゼ(PHD)を制御するのがシュウ酸濃度なのか、実際に存在するTRPA1の量そのものなのかを評価するため、タンパク質レベルでのTRPA1の存在量も調べた。その結果、3つの条件下で統計的に有意な差は見られなかった(図S12)。次に、様々な動物種において、熱ストレス時にTRPA-1の発現に影響を与え、共益的に作用することが知られているHIF1αのタンパク質量を調べた(Bairdら、2006、Brugalettaら、2022、Hatanoら、2012、Klumpenら、2017、Zhangら、2023)。その結果、熱ストレス時にHIF1αのタンパク質レベルが高くなる傾向が確認された。以上述べたことから、今回のデータでは図7cのカスケードが想定される。TRPA-1の反応性を調節する酵素として知られているPHDが、シュウ酸/シュウ酸塩の減少によって制御されている可能性が予想される(緑色の矢印で示したカスケード)。この場合、温度感受性は高くなく、寒さには弱くなる(図7bおよび7c)。本研究の観察結果から、熱ストレス条件下でのW. coagulans投与による効果は、同様のカスケードに従っているようである。参考データとして、本実験では通常条件下でブロイラーのシュウ酸/シュウ酸塩の肝濃度が上昇した(図4)。通常条件下で低温感受性が高くても、高温条件下ではないため、成長阻害作用は生じないと考えられた(表1)。これらの仮説に基づき、これらの反応に関連する因子を評価した。様々なマーカーが豊富に存在し、検出キットの種類も豊富なマウスと異なり、ニワトリ組織におけるPHDの生理反応を定量的に評価することは困難である。そこで、ニワトリ由来の分子と交差活性を持つ抗体を用いて、以下のマーカーを調べた。PHDはHIF1αの発現を促進し(Kikuchi et al., 2014)(図7c)、HIF1αのタンパク質レベルは一般にTRPA1の発現誘導に関与しているので(Baird et al., 2006, Brugaletta et al、 2022, Hatano et al., 2012, Klumpen et al., 2017, Zhang et al., 2023)(図7c)上述したように、反応を評価するために、間接的なマーカー分子としてのHIF1αのタンパク質量を調べた(図S12)。通常条件と熱ストレス条件では異なる傾向が認められたが(p = 0.1211)、熱ストレス条件では2群間の差は認められなかった。そこで、別の視点に着目した。TRPA-1の存在は昆虫からヒトまで共通であり(Kurganov et al., 2017)、この視点はモデル動物以外の視点も加味している可能性がある。この反応性の観点からは、抗酸化物質や過酸化物は、活性酸素種(ROS)そのものが考慮される重要な代謝産物である(Miyake et al.) 興味深いことに、これらの実験条件下における糞便および肝臓中のアスコルビン酸濃度は、熱ストレス試験群(H_coagulans)で高い傾向にあった(p < 0.1)(図S13a)。しかしながら、過酸化脂質の安定な指標であるマロンジアルデヒドのレベル(Miyamoto et al. これらのデータから、本研究のブロイラーモデルにおけるカスケードは、以前報告されたような低酸素ストレス条件(Miyake et al.) 本研究の条件下では、熱ストレス試験群(H_coagulans)は、メチルマロン酸(Fernandesら、2011、Pillaiら、2019、Rosenbergら、1968)(p = 0.0526)およびパラキサンチン(Gressnerら、2009)(p = 0.0526)の減少傾向(p < 0.1)を示した(図S13a)ことから、肝機能抑制作用があると考えられる。このような変化をもたらすもう一つの詳細な作用機序は、今回の研究では見つからなかった。少なくとも、W. coagulansの曝露は、耐熱性下での腸-肝臓相互作用のこのような特徴的な代謝カスケードに影響を及ぼすという仮説が立てられた。
腸内細菌群の構造方程式と因果推論から、フラボニフラクトール属、シュードモナス属、ロンブスティア属からなるネットワークとして強い関係が示され、単一の要因ではなく、グループ全体としての重要性が示された。ネットワーク内の個々の細菌の特徴は以下のように評価された。Flavonifractor属は、低いレベルではあるが、熱試験群(H_coagulans)で増加しているように見え(図5bおよびS9a)、有意ではない(p = 0.2629)ものの、因果推論は必須因子として位置づけられなかった(図5d)。近年、有益菌候補としてFlavonifractor属に属する菌株が注目されている(Mikami et al.) また、Romboustia属も、有意ではないが、熱試験群(H_coagulans)で増加しているように見えた(図5bおよびS9a)(p = 0.0855)。細菌解析の結果、体重増加に関連する因子として因果関係が推論された(値=6.53)(図5cおよび5d)。Romboustia属に属する菌株は栄養豊富な環境に適応している(Gerritsen et al.) 一方、Pseudomonas属は、低レベルではあるが、熱試験群(H_coagulans)では有意ではないものの減少しているように見えた(図5bおよびS9a)(p = 0.1732)。Pseudomonas属は、環境中で様々な機能を果たす以下の細菌として知られている: P.aeruginosaは動物の日和見病原細菌(Hassetta et al., 2002)、P.syringae(Hirano & Upper, 2000)は植物病害の原因菌、P.putidaは生物農薬(Espinosa-Urgel et al., 2000)や環境汚染物質の分解菌(Gomes et al., 2005)、P.fluorescens(Haas & Defago, 2005)は植物成長促進菌である。本研究では、これらのグループの効果を正確に把握することは不可能である。少なくとも、W. coagulansが熱ストレス下のブロイラー鶏の糞便中の細菌ネットワークをグループとして制御できることは計算上確認されており、この条件下でW. coagulansによる熱耐性が生じたことは事実として受け止める必要がある。
まとめとして、W. coagulansが耐熱性に関与する可能性のあるメカニズムを図7cに示す。W. coagulansは、緑色の矢印で示したように、腸内細菌叢を変化させることによって、腸内代謝産物の挙動に影響を与える。腸内細菌であるシュードモナスの存在は、一般的にピリドキサール濃度に影響を与える酵素を増強し、その減少に影響を与える。第二に、ピリドキサールの増加は肝シュウ酸/シュウ酸塩の濃度に影響し、その結果PHDの水酸化が阻害されない可能性がある。したがって、TRPA1による寒冷感受性は増加しない可能性がある。さらに、これらのメカニズムは、熱ストレスやその他の抗酸化ストレスから、薬剤を使わずにプロバイオティクス由来で緩和される可能性を示唆しているかもしれない。今後の実験ステップとして、ピリドキサール自体の経口投与と、熱ストレス条件下での肝臓のシュウ酸/シュウ酸塩濃度との関係を評価する研究により、本研究の構造式に基づく予測が実証されることが期待される。さらに、熱ストレス応答に関するチェックポイントの検証も必要であろう。このように、腸内常在細菌叢の特性との関係を詳細に検討することで、今後の基礎となる新たな暑熱耐性のメカニズムとその対処法を得ることができるであろう。
方法
4.1. ブロイラーの管理
すべての動物実験は千葉大学動物実験施設管理使用委員会(DOU27-131, DOU28-157, DOU30-331, DOU1-335)の承認を受け、千葉県畜産試験場ガイドライン(Ito et al.) ブロイラー雛(チャンキー)は、バッテリーブルーダー(幅85cm×奥行き85cm×高さ25cm)内で飼料を自由摂取可能な飼料(CP, 22 %; ME, 3,100 kcal/kg)を用いて10日間慣行飼育し、その後、10日目以降21日目まで3群に分離した。無作為に選択したヒナを維持し(1ケージあたりn = 3)、制限された給餌エリア(幅22 cm x 奥行き44 cm x 高さ45 cm)に合わせてヒナを選択した。21日目から49日目まで、各ひよこは別の制限給餌エリア(ケージあたりn = 1)(幅44 cm x 奥行き56 cm x 高さ65 cm)に維持された。すべてのヒナには、孵化後6日目まで抗生物質としてテトラサイクリン系に属するドキシサイクリンKS(共立製薬株式会社)を50mg/L、水中に経口投与した。W. coagulans SANK70258 (Lacris-10)(三菱化学(株))は、最初の10日間は自由飲水により最終濃度として少なくとも105 CFU/mlの濃度で飲料水に投与し、その後は飼料添加物として0.005 %の濃度で投与した(少なくとも105 CFU/g)。糞および肝臓はすべて、解剖時(飼育終了後 49 日目)の午前 8 時から午後 2 時の間に採取した。
4.2. HPLC用糞便サンプルの調製
得られた糞便サンプルはすべて-20℃で輸送し、その後分析まで-60℃~-80℃で保存した。糞便サンプルは、製造業者のプロトコール(Tonda et al. 簡単に説明すると、ニワトリの糞便(200~400 mg)を9倍量のMilli-Q水と10分間混合した。15,000rpmで遠心した後、上清を0.45μmのMillex-HAフィルター(SLHA033SS)(Millipore、米国)でろ過した。最後に、ろ過した溶液をHPLC装置のHPLC分析用に調製した。酢酸、プロピオン酸、酪酸および乳酸の濃度を測定するために、イオン排除カラム(Shim-pack SCR-102H)と電気伝導度検出器(CDD-10AVP)(島津製作所)を備えたHPLC Prominence装置(島津製作所)を用いて、凍結新鮮糞便サンプルの含有量を調べた。
測定条件は次のように調整した:移動相、5 mM p-トルエンスルホン酸;緩衝液、5 mM p-トルエンスルホン酸、20 mM Bis-Tris、0.2 mM EDTA-4H;温度、40 ℃;流速、0.8 mL/分。
4.3. GC/MS/MS用糞便サンプルの調製
水溶性代謝物を測定するために、凍結乾燥した糞便サンプルを処理した。GC/MS/MS装置とGCMS-TQ8030トリプル四重極型質量分析計(島津製作所、日本)を用い、以前に記載された方法(Sato et al. 鶏排泄物は消化されにくい傾向があるため、いくつかの前処理工程を施した。凍結乾燥した糞便を、Micro Smash MS-100(Tomy Seiko, Japan)を用いてジルコニアビーズで破砕した。次に、5 mgの糞便サンプルを、15 μLの内部標準物質(1 mM 2-イソプロピルマレイン酸)を含む150 μLのMilli-Q水に懸濁し、150 μLのメタノールと60 μLのクロロホルムを加えた。これらの溶液を混合し、37℃で30分間、1200rpmで振とうしながらインキュベートした。16,000g、5分間、室温で遠心した後、上清(水層)250μLを新しいチューブに移し、クロロホルム60μLを加えた。混合後、溶液を37℃で30分間、1200rpmで振盪しながらインキュベートした。各溶液を室温で16,000×g、5分間遠心し、上清250μLを新しいチューブに移し、Milli-Q水200μLを加えた。溶液を16,000 gで5分間、室温で遠心分離し、上清125 μLを真空エバポレーターシステム(CentriVap Centrifugal Vacuum Concentrator, Labconco, USA)を用いて40℃で20分間蒸発乾固し、凍結乾燥機(Taitec, Japan)を用いて凍結乾燥した。乾燥抽出物は、その後の前処理を測定直前に行うまで-80℃で保存した。乾燥抽出物をまず、ピリジンに溶解した20 mg/mL メトキシアミン塩酸塩(Sigma-Aldrich、米国)40 μLでメトキシル化した。その後、20μLのN-methyl-N-trimethylsilyl-trifluoroacetamide (MSTFA) (GL Science, Japan)を用いて、37℃で30分間、振盪しながらインキュベートした。誘導体化後、室温で16,000×g、5分間遠心し、上清をガラスバイアルに移し、GC-MS/MS測定を行った。GC-MS/MS分析は,島津GCMS-TQ8030トリプル四重極質量分析計(島津製作所)を用い,キャピラリーカラム(BPX5;SGEアナリティカルサイエンス社製)を用いて行った。GCオーブンは以下のようにプログラムされた: 60 °Cで2分間保持した後、330 °Cまで昇温し(15 °C/分)、最後に330 °Cで3.45分間保持した。GCは39 cm/sの一定線速度モードで行った。検出器の温度は200 °C、インジェクターの温度は250 °Cであった。注入量は1μL、スプリット比は1:30とした。データ処理はLabSolutions Insight(Shimadzu, Japan)を用いて行った。
4.4. エンリッチメント解析
MetaboAnalyst 5.0 (https://www.metaboanalyst.ca)(Xia et al., 2009, Xia and Wishart, 2011a, Xia and Wishart, 2011b)を用いて、検出された特徴的な代謝物(p < 0.1; 熱ストレス対照群のデータに対して1.5倍変化)のパスウェイ解析および濃縮解析を行った。
4.5. 糞便サンプルからのDNA調製
糞便から全DNAを既報(Kim et al. 凍結乾燥した糞便約100 mgをMicro Smash MS-100(Tomy Seiko, Tokyo, Japan)を用いてジルコニアビーズで破砕し、10 mM Tris-HCl(pH 8.0)、1 mM EDTA(TE)を含む溶液600 μLに懸濁した。475μLの糞便懸濁液を1.5mLの微量遠心チューブに移した。この懸濁液を15 mg/mLのリゾチームとともに37℃で1時間インキュベートした。次に、アクロモペプチダーゼ(富士フイルム和光純薬、大阪、日本)を終濃度2,000ユニット/mLで加え、さらに37℃で30分間インキュベートした。次いで、プロテイナーゼKおよびドデシル硫酸ナトリウムを、それぞれ最終濃度1 mg/mLおよび1 %で25.7 μL添加し、混合溶液を55℃で1時間インキュベートした。得られた溶解液をフェノール/クロロホルム/イソアミルアルコールで処理し、DNAをエタノールで沈殿させた。DNAペレットを70%エタノールで洗浄、乾燥し、300μLのTEに溶解した。次に、RNaseを最終濃度0.1 mg/mLで加え、溶液を37℃で30分間インキュベートした。その後、2.5M NaClを含む20%ポリエチレングリコール6000溶液を等量加えた。氷上で30分間インキュベートした後、遠心分離でDNAをペレット化し、水洗した後、TEに溶解した。
4.6. 16S rRNA シークエンシング解析
16S rRNA遺伝子の解析は、以前の報告(Kim et al、 2013). PCR反応ミックスは、DNA鋳型4 µL、ExTaqポリメラーゼ0.5 µL(タカラバイオ株式会社、日本、草津)、ExTaqバッファー5 µL、dNTPs 5 µL、フォワードおよびリバースプライマー各10 pmolからなり、DNAを含まない水で全量を50 µLに調整した。アンプリコンは、98 °Cで30秒、55 °Cで45秒、72 °Cで2分のサイクルを30回行い、AMPure XP(Beckman Coulter Inc.) マルチプレックスアンプリコンシークエンシングは、以下に記載する条件で行った。製造元の指示に従い、サンプルは454 GS FLX/Juniorを用いて塩基配列を決定した。平均品質値が25未満のリード、または両方のユニバーサルプライマーに正確に一致しないリードはフィルターで取り除かれた。最後に、フィルターされたリードをさらなる解析に使用した。次に、これらのリードを平均品質値に従ってソートし、UCLUST (https://www.drive5.com/)を用いて操作上の分類単位(OTU)にグループ化した。配列同一性の閾値は454データで96 %であった。生成されたOTUの代表配列は、GLSEARCHプログラムを用いて上記のデータベースとの相同性検索を行い、分類学的割り当てを行った。門、属、種レベルでの分類には、それぞれ70 %、94 %、97 %の配列類似度閾値を適用した。α-多様性の指標、すなわち群集の豊かさ(Chao1)と多様性(Shannon and Simpson)を計算し、加重および非加重主座標分析(PCoA)を用いたUniFrac分析によりβ-多様性の指標も推定した。Alla16S rRNA遺伝子のデータセットはGenBank Sequence Read Archiveデータベースに寄託された。
4.7. 相関分析
糞便代謝物間のピアソン相関係数をRソフトウェアで計算した後、相関ヒートマップを構築することにより、支配的および/または特徴的な糞便代謝物の相対値を可視化した(Ito et al.)
4.8. 機械学習
メタボロームデータの特徴因子は、教師ありML、ランダムフォレスト(Breiman, 2001)、バギング(ブートストラップ集計)を用いたMLの1つ(Miyamoto & Kikuchi, 2023)、エクストリームグラジェントブースティングを用いたMLの1つ(Miyamoto & Kikuchi, 2023)であるXGBoost(Chen & Guestrin, 2016)によってそれぞれ抽出した。オブザベーションの数が少なくても,ブートストラップ法を用いたランダム・サンプリングによって一定のサブサンプルを生成でき,選択された説明変数は,最良の分類因子とそのしきい値を計算するために使用できる.本研究では、このML特性の利点を活かし、限られた実験条件下で、細菌と代謝産物の特徴的な一群を抽出した。特徴的な成分は、Rソフトウェアパッケージ "randomForest" (https://cran.r-project.org/web/packages/randomForest/index.html)と "xgboost" (https://cran.r-project.org/web/packages/xgboost/index.html)を用いて選択した。
4.9. 動物種間の構造方程式の推定
確証的因子分析(CFA)のための構造方程式モデリング(SEM)は、主に有向非循環グラフ(DAG)の数が多い成分群について実施した。SEMは、Rソフトウェアのパッケージ "lavaan"(Rosseel, 2012, Rosseel et al., 2021)を用いて、既述(Miyamoto et al.) 解析コードはウェブサイト(https://lavaan.ugent.be)を参照した。CFAには仮説が必要であるため、関連解析で選択されたグループを代謝物と微生物叢の潜在構成因子の因子として利用した。仮説としてのモデルは、ブートストラップ(n = 1000)を用いた最尤(ML)パラメータ推定を用いて、関数「lavaan」と「sem」によって統計的に推定した。モデルの適合性は、カイ二乗p値(p > 0.05、有意ではない)、比較適合指数(CFI/cfi)(>0.9)、Tucker-Lewis指数(TLI/tli)(>0.9)、適合度指数(GFI/gfi)(>0. 95)、修正適合度指数(AGFI/agfi)(>0.90)、近似二乗平均平方根誤差(RMSEA/rmsea)(<0.05)、標準化平均残差(SRMR/srmr)(<0.08)が、良好なモデル適合の指標である(Hooper et al、 2008). 最適な構造式を選択する際、上記の条件を満たす場合、GFIとAGFIの値の差が少ない式を採用した。次に、候補モデルが多い場合は、赤池情報量規準(AIC)の値が小さいモデルを採用した。良いモデルのパス図は、Rソフトウェアのパッケージ "semPlot"(Epskamp et al.)
また、動物種を超えた構造的普遍性を評価するために、機械学習により選択された因子群に対する統計的因果推論として、線形非ガウス非周期モデルおよび因果関係推定のための非ガウス独立成分分析であるDirectLiNGAM(Shimizu et al. DirectLiNGAMは、既述(Miyamoto et al., 2022, Miyamoto et al., 2022, Okada et al., 2023)の通り、ウェブサイト(https://github.com/cdt15/lingam)上のPythonコードで構築した。本研究では、Pythonバージョン3.11.2上で以下のパッケージを使用した: Cartopy 0.21.1、graphviz 0.20.1、libarchive 0.4.7、lingam 1.7.1、numpy 1.23.4、pandas 1.5.3、pygam0.8.0。DirectLiNGAMで計算したデータに基づくDAGをGephi(バージョン0.9.2)で可視化(Force Atlas with Noverlap)した(https://gephi.org)。モジュラリティクラスはGephiによって自動的に判別された。
すべての構造方程式は、成長に関するデータを除いた生データを用いて分析された。成長に関しては、2値化によってグループ間の有意差を表し、欠損値の影響を排除した(Datafinal_Lac.xlsx; https://github.com/hmiyamoto2000/Paper_Heat_stress を参照)。
4.10. コードダイアグラムによる俯瞰分析
動物種ごとの影響度の可視化は、Rソフトウェアのパッケージ "circlize "を用いたChord diagramによって行った。実験条件による差異への影響を避けるため、すべてのデータは同一成分のデータの中央値(M)をもとに算出し、0(<M)と1(>M)にソートした。
4.11. イムノブロット
50mgの組織を、プロテアーゼ阻害剤:10μL/mLのPMSF(SigmaP7626)(Sigma, USA)、10μL/mLのオルトバナジン酸ナトリウム(Sigma S6508)(Sigma, USA)、および20μL/mLのプロテアーゼ阻害剤カクテル(Thermo Scientific Halt Protease Inhibitor Single-Use Cocktail #78430 )(Thermo Scientific, USA)を含む氷冷PBS-T 150μL中に移した。ソニケーター Ultrasonic processor XL2020 (MISONIX, USA)を用いて氷上でホモジナイズし、4℃、10,000×gで5分間遠心して上清を回収した。上清のタンパク質濃度は、Thermo Scientific Pierce BCA Protein Assay kit #23227 (Thermo Scientific, USA)を用いて測定した。
粗タンパク質溶液を7.5 %ドデシル硫酸ナトリウムポリアクリルアミドゲル上で電気泳動により分離し、PVDF膜(GE Healthcare Life science, Sweden)に転写した。膜はPVDFブロッキング試薬(東洋紡、日本)で室温で1時間ブロッキングした。一次抗体は、1:1500に希釈したウサギポリクローナルTRPA1抗体(19124-1-AP)(proteintech group, Japan)または1:1500に希釈したウサギポリクローナルHIF-1α抗体(bs-20399R)(Bioss andibodies, USA)を用い、二次抗体は1:12000に希釈した西洋ワサビペルオキシダーゼ標識ヤギ抗ウサギIgG抗体(Cell Signaling Technology, USA)を用いた。GAPHDHは、1:10000に希釈したウサギポリクローナルGAPDH抗体(10494-1-AP) (proteintech group, Japan)を用いて検出した。イムノブロットは、一次抗体を4℃で一晩、ペルオキシダーゼ標識二次抗体を室温で1時間インキュベートした後、Western Lightning TM ECL Pro(Perkin-Elmer Life Sciences, USA)を用いてタンパク質シグナルを検出した。化学発光スキャナーC-DiGiT(スクラム、日本)とImage Studioを使用して、タンパク質のシグナルを測定した。
4.12. マロンジアルデヒド濃度の分析
肝臓および白色脂肪細胞組織中のマロンジアルデヒド(MDA)濃度は、MDA測定キットBioxytech MDA-586(Percipio Bioscience, CA, USA)を用いて、既述のように製造者のプロトコールに従って測定した(Miyamoto et al.) すなわち、50 mgの組織をBHT含有PBS-T(0.05 % tween 20含有)中で超音波処理装置Ultrasonic processor XL2020 (MISONIX, USA)を用いてホモジナイズし、45℃で60分間インキュベートした後、10,000 × gで10分間遠心した。その後、上清を586 nmで測定した。
4.13. 統計分析
大腸糞便細菌叢および代謝物濃度のデータは以下のように解析した:ガウス分布を評価するためにShapiro-Wilk検定を用いた。さらに、Bartlett 検定を用いて等分散性を評価し、パラメトリック解析とノンパラメトリック解析を選択した。パラメトリック分析としては、ANOVAに続いてTukeyのpost hoc検定が用いられ、ノンパラメトリック分析としては、Kruskal-Wallisの一元配置分散分析に続いてSteel-Dwass検定が行われた。これらの計算データは、Rソフトウェア(バージョン3.6.2、4.0.5、4.2.2)、GraphPad Prismソフトウェア(バージョン9.1.2、9.2.0)(GraphPad Software, USA)、Microsoft Office(バージョン16.66.1)を用いて作成した。パスウェイ解析のデータは、MetaboAnalyst 5.0 (Xia et al., 2009, Xia and Wishart, 2011a, Xia and Wishart, 2011b)を用いて計算し、可視化した。パスウェイ解析は、低濃度(≥0.1μmol/g; p < 0.05)ではなく、統計的に有意な代謝物を対象として行い、顕著な特徴を把握した。p<0.05で有意、0.05≦p<0.20で傾向とした。データは平均値±SEで示した。
CRediT著者貢献声明
伊藤佳代.執筆 - 原案, 資料, プロジェクト管理, 調査, データキュレーション, 概念化. 宮本博邦: 執筆-校閲・編集、執筆-原案、可視化、検証、監修、ソフトウェア、資源、プロジェクト管理、方法論、調査、資金獲得、形式分析、データキュレーション、概念化。松浦真紀子:プロジェクト管理、調査、形式分析。石井千歳 プロジェクト管理、調査。中西由美子:調査、形式分析、データキュレーション。須田渉 調査、形式分析、データキュレーション。佐藤隆史: 形式分析。本田冬子:プロジェクト管理。黒谷淳: ソフトウェア、方法論、調査、データキュレーション。辻直子: 調査、形式分析、データキュレーション。中熊輝乃:調査。進藤千絵: リソース、方法論、調査。伊藤俊之 プロジェクト管理。道端龍之介:執筆(校閲・編集)、リソース。山田亮一:執筆-校閲・編集、監修、リソース。高橋洋介:監修、リソース、調査、構想。児玉裕明: 執筆(校閲・編集)、監修、リソース、方法論、調査。菊池純: 執筆(校閲・編集)、ビジュアライゼーション、監修、ソフトウェア、方法論、調査。大野裕史: 執筆-校閲・編集、監修、資料、方法論、調査。
利益相反宣言
著者らは、本論文で報告された研究に影響を及ぼすと思われる競合する金銭的利益や個人的関係がないことを宣言する。
謝辞
本研究の一部は、経済産業省「ものづくり基盤整備事業」に基づく千葉県中小企業団体中央会の助成金(助成番号25121200402)の支援を受けた。岩田祥子氏(千葉大学)には、飼料のサンプリングおよび調製にご協力いただいた。また、特別な助言をいただいた葛城俊哉博士、古賀裕久氏、渡辺裕子氏、柏慶太氏、前田隆之氏(三菱化学)、スタッフ・教員の特別調整をしていただいた脇雅之氏、高橋啓二氏、奥田裕子氏(千葉大学)に感謝する。また、高橋啓二氏、岡田裕子氏、藤崎忠彦氏、高橋康弘氏、多田義明氏、後藤晴一氏(千葉県畜産試験場)には、飼育施設の管理、サンプリング、飼料の調製について特別な助言をいただいた。また、GCMS/MS実験の技術支援をしていただいた朝倉美紀氏、伊藤あゆみ氏(理研IMS)に感謝する。
倫理的声明
本研究におけるすべての動物実験の関与に関する倫理的承認は、千葉大学Institutional Animal Care and Use Committeeによって行われた(参照番号:DOU27-131, DOU28-157, DOU30-331, DOU1-335)。
データの利用可能性
細菌のV1-V2 16S rRNAデータのRawファイルは、NCBI Bio-Projectアクセッション番号PRJDB10703 (PSUB013670)で、日本DNAデータバンク(DDBJ)に寄託されている(BioSampleアクセッション番号: SAMD00261459-SAMD00261461, SAMD00253848-SAMD00253854).
付録A. 補足データ
本論文の補足データです:
ダウンロード ダウンロード:PDFファイル(1MB)
補足データ1.
参考文献
Ara et al.
K. 荒 浩二, 目黒 聡, 長谷 崇, 時光 郁夫, 尾辻 浩二, 河合 聡, ..., 飯野 均
有胞子性乳酸菌(Bacillus coagulans SANK 70258)投与がヒトおよびラットの腸内環境、排便回数、糞便性状および皮膚性状に及ぼす影響
健康と病気の微生物生態学, 14 (1) (2009), pp.4-13, 10.1080/089106002760002694
グーグル・スカラー
バガスら、2019
M. バガス、G.クリシュナン、C.デバラジ、V.P.ラシャモール、P.プラニャ、A.M.リース、V.セジャン
乳牛の免疫系に対する暑熱ストレスの影響: レビュー
Research in Veterinary Science, 126 (2019), 94-102頁, 10.1016/j.rvsc.2019.08.011
PDFで記事を見るScopusで記事を見るGoogle Scholar
ベアードら、2006
N.A.ベアード、D.W.ターンブル、E.A.ジョンソン
低酸素状態における熱ショック経路の誘導には、低酸素誘導性因子-1による熱ショック因子の制御が必要である。
Journal of Biological Chemistry, 281 (50) (2006), 38675-38681頁, 10.1074/jbc.M608013200
PDFで記事を見るScopusで記事を見るGoogle Scholar
ベッカーら、2021年
C.A.ベッカー、A.アガラリ、M.マルフザマン、A.E.ストーン
機械学習技術を用いた乳牛の暑熱ストレス予測
Journal of Dairy Science, 104 (1) (2021), pp.501-524, 10.3168/jds.2020-18653
PDFを見る記事を見るScopusGoogle Scholarで見る
ブレイマン, 2001
L. ブレイマン
ランダムフォレスト
機械学習, 45 (2001), pp.
Google Scholar
ブラウン他, 2012
S.D. Brown, S.M. Utturkar, D.M. Klingeman, C.M. Johnson, S.L. Martin, M.L. Land, ..., D.A. Pelletier
ポピュラス・デルトイデスの根圏および終末圏から分離されたシュードモナス属細菌21種と多様な細菌19種のゲノム配列
Journal of Bacteriology, 194 (21) (2012), pp.5991-5993, 10.1128/JB.01243-12
スコープで見るGoogle Scholar
ブルガレッタら、2022年
G.ブルガレッタ、E.グリーン、A.ラムザー、C.W.メイナード、T.W.タブラー、F.シリ、...、S.ドリディ
ジャングル家禽および3つのブロイラー系研究系統における視床下部の酸素ホメオスタシスと炎症状態に対する周期的熱ストレスの影響
Front Vet Sci, 9 (2022), Article 905225, 10.3389/fvets.2022.905225
スコープで見るGoogle Scholar
ブルガレッタら、2022年
G. Brugaletta、J.R. Teyssier、S.J. Rochell、S. Dridi、F. Sirri
鶏の熱ストレスに関する総説。第1部:生理学と腸内健康への洞察
Frontiers in Physiology, 13 (2022), Article 934381, 10.3389/fphys.2022.934381
スコープで見るGoogle Scholar
チェリーニら、2009
B. チェリーニ、R.モンティオーリ、A.パイアルディーニ、A.ロレンツェット、C.B.ヴォルタットルニ
ヒト肝ペルオキシソームアラニンのマイナーアレルの相乗作用に関する分子的洞察: グリオキシル酸アミノトランスフェラーゼのマイナーアレルとF152I変異との相互作用に関する分子的洞察
Journal of Biological Chemistry, 284 (13) (2009), pp.8349-8358, 10.1074/jbc.M808965200
PDFで記事を見るScopusで記事を見るGoogle Scholarで記事を見る
チェンとゲストリン, 2016
T. チェン、C. ゲストラン
XGBoost
第22回ACM SIGKDD知識発見とデータマイニング国際会議予稿集(2016), pp.785-794
CrossRefGoogle Scholar
クレイマーら、2022
M.N.クレイマー、D.ギャニオン、O.ライタノ、C.G.クランドール
健康、疾病、傷害における熱ストレス下でのヒトの体温調節
Physiological Reviews, 102 (4) (2022), pp.1907-1989, 10.1152/physrev.00047.2021
Scopusで見るGoogle Scholar
クレイマーら、2018
T.A.クレイマー、H.W.キム、Y.チャオ、W.ワン、H.W.チェン、Y.H.B.キム
慢性熱ストレスに暴露したブロイラーの胸筋の肉質特性に及ぼすプロバイオティクス(枯草菌)補給の効果
Poultry Science, 97 (9) (2018), pp.3358-3368, 10.3382/ps/pey176
PDFで記事を見るScopusで記事を見るGoogle Scholar
Dauria et al.
B.D. Dauria, A. Sigdel, J. Petrini, P.P. Boscollo, F. Pilonetto, M. Salvian, ..., G.B. Mourao
ブラジル産ホルスタイン牛の乳脂肪酸に対する暑熱ストレスの遺伝的影響
Journal of Dairy Science, 105 (4) (2022), pp.3296-3305, 10.3168/jds.2021-20914
PDFで記事を見るScopusで記事を見るGoogle Scholarで記事を見る
デルワースら、1999
T.L.デルワース、J.D.マールマン、T.R.クヌートソン
CO2による地球温暖化に伴う暑さ指数の変化
Chimatic Change, 43 (1999), pp.369-386, 10.1023/A:1005463917086
スコープで見るGoogle Scholar
エプスカンプら、2019
Epskamp, S., Stuber, S., Nak, J., Veenman, M., & Jorgensen, T. D. (2019). 様々なSEMパッケージの出力のパス図と視覚的分析Rパッケージバージョン1.1.2.
Google Scholar
エスピノサ・ウルゲルら、2000年
M. エスピノサ・ウルゲル、A.サルディオ、J.L.ラモス
Pseudomonas putidaの種子付着に関与する機能の遺伝学的解析
Journal of Bacteriology, 182 (2000), pp.2363-2369, 10.1128/JB.182.9.2363-2369.2000
スコープで見るGoogle Scholar
フェルナンデスら、2011
C.G. Fernandes, C.G. Borges, B. Seminotti, A.U. Amaral, L.A. Knebel, P. Eichler, ..., M. Wajner
メチルマロン酸が若齢ラットの神経終末および線条体において酸化的損傷を誘発し、抗酸化防御を損なうという実験的証拠
細胞分子神経生物学, 31 (5) (2011), pp.775-785, 10.1007/s10571-011-9675-4
スコープで見るGoogle Scholar
ガティラムら、1987
P. Gathiram, M.T. Wells, J.G. Brock-Utne, B.C. Wessels, S.L. Gaffin
熱ストレスにおける非吸収性抗生物質による内毒素血症の予防
Journal of Clinical Pathology, 40 (1987), pp.1344-1368, 10.1136/jcp.40.11.1364
グーグル奨学生
ゲリッツェンら、2017年
J. Gerritsen, B. Hornung, B. Renckens, S. van Hijum, V.A.P. Martins Dos Santos, G.T. Rijkers, ..., H. Smidt
Romboutsia ilealis CRIB(T)のゲノムおよび機能解析から、小腸への適応が明らかになった
PeerJ, 5 (2017), p. e3698
CrossRefView in ScopusGoogle Scholar
Goelら、2021年
A. Goel, C.M. Ncho, Y.H. Choi
熱ストレスによるニワトリの遺伝子発現制御
Journal of Animal Science and Biotechnology, 12 (1) (2021), p. 11, 10.1186/s40104-020-00523-5
ScopusGoogle Scholarで見る
ゴメスら, 2005
N.C.ゴメス、I.A.コシェレバ、W.R.エイブラハム、K.スモーラ
接種菌株Pseudomonas putida KT2442(pNF142)およびナフタレン汚染が土壌細菌群集に及ぼす影響
FEMS Microbiology Ecology, 54 (1) (2005), pp.21-33, 10.1016/j.femsec.2005.02.005
PDFを見る記事を見るScopusで見るGoogle Scholarで見る
グレスナーら, 2009
O.A.グレスナー、B.ラーメ、M.シルシェク、A.M.グレスナー
肝実質細胞における結合組織成長因子発現の最も強力なカフェイン由来阻害剤としてのパラキサンチンの同定
Liver International, 29 (6) (2009), pp.886-897, 10.1111/j.1478-3231.2009.01987.x
スコープで見るGoogle Scholar
ハースとデファゴ, 2005
D. Haas, G. Defago
蛍光性シュードモナドによる土壌伝染性病原菌の生物学的防除
Nature Reviews. 微生物学, 3 (4) (2005), 307-319頁, 10.1038/nrmicro1129
スコープで見るGoogle Scholar
ハセッタら、2002年
D.J. Hassetta、J. Cuppolettib、B. Trapnellc、S.V. Lymar、J.J. Rowe、S.S. Yoon、...、U.A. Ochsner
慢性的に感染した嚢胞性線維症気道における緑膿菌バイオフィルムによる嫌気性代謝とクオラムセンシング: 抗生物質治療戦略と創薬標的の再考
Advanced Drug Delivery Reviews, 54 (2002), pp.1425-1443, 10.1016/s0169-409x(02)00152-7
Google Scholar
波多野ほか, 2012
N. 波多野, 伊藤靖之, 鈴木秀樹, 村木靖之, 林秀樹, 小野崎和彦, ..., 村木和彦
低酸素誘導因子-1α(HIF1α)は、低酸素応答要素様モチーフを介して一過性受容体ポテンシャルアンキリンリピート1(TRPA1)遺伝子の発現をスイッチし、サイトカイン放出を調節する
Journal of Biological Chemistry, 287 (38) (2012), 31962-31972頁, 10.1074/jbc.M112.361139
PDFで記事を表示Scopusで記事を表示Google Scholar
Heら、2020
Y. He、C. Maltecca、F. Tiezzi、E.L. Soto、W.L. Flowers
トランスクリプトーム解析により、ブタの暑熱耐性の基礎となる遺伝子と共発現ネットワークが同定された。
BMC Genetics, 21 (1) (2020), p. 44, 10.1186/s12863-020-00852-4
PDFを見る記事を見るGoogle Scholar
平野・上, 2000
平野修一、C.D.アッパー
葉の生態系における細菌-病原体、氷核、着生植物であるシュードモナス・シリンゲを中心に
微生物学・分子生物学レビュー, 64 (3) (2000), pp.624-653, 10.1128/mmbr.64.3.624-653.2000
ScopusGoogle Scholarで見る
ホロウェイら、2016
K.A.ホロウェイ、L.ロゼッラ、D.ヘンリー
WHO必須医薬品政策が抗生物質の不適切使用に与える影響
PLoS One1, 11 (3) (2016), p. e0152020
CrossRefView in ScopusGoogle Scholar
フーパーら、2008年
D. フーパー、J.コフラン、M.R.ミューレン
構造方程式モデリング: モデル適合を決定するためのガイドライン
電子ジャーナルビジネス研究法, 6 (2008), pp.53-60
スコープで見るGoogle Scholar
石田ら, 2022
S. 石田、大澤、山口修一、山崎哲也、萩谷和夫
日本におけるホルスタイン牛の暑熱耐性の遺伝学的評価に適した暑熱ストレス指標
畜産学雑誌, 93 (1) (2022), p. e13688
スコープで見るGoogle Scholar
伊藤ら, 2022
K. 伊藤紘一, 宮本博之, 松浦正樹, 石井千尋, 坪井明, 辻直樹, ..., 大野均
ブロイラー用耐熱性乳酸菌Weizmannia coagulans SANK70258の特性評価のための非侵襲的糞便代謝プロファイリング
バイオサイエンス・バイオエンジニアリング, 134 (2) (2022), pp.105-115, 10.1016/j.jbiosc.2022.05.006
PDFで記事を見るScopusで記事を見るGoogle Scholar
伊藤ら、2019
K. 伊藤紘一, 八木健一, 本田文彦, 岡田裕之, 高橋和樹, 脇雅史
ブロイラーの肥育ステージに応じた的確な栄養管理による暑熱適応技術
千葉県畜産研究センター紀要, 19 (2019), pp.
グーグル・スカラー
カーンら, 2019
A.Z.カーン、I.U.カーン、S.カーン、S.アフザル、M.ハミド、M.タリク、...、R.リュー
セレン強化プロバイオティクスは、熱ストレス条件下におけるブロイラーの炎症性サイトカインおよび抗酸化能を制御することにより、肝保護を改善する。
Journal of Advanced Veterinary Research, 6 (3) (2019), pp.355-361, 10.5455/javar.2019.f354
ScopusGoogle Scholarで見る
菊池ら、2014
D. 菊池大介, 南島義明, 中山和彦
プロリルヒドロキシラーゼPHD3はピルビン酸デヒドロゲナーゼ(PDH)-E1βと相互作用し、細胞内PDH活性を制御する
生化学・生物物理研究通信, 451 (2) (2014), pp.288-294, 10.1016/j.bbrc.2014.07.114
PDFで記事を見るScopusで記事を見るGoogle Scholar
キムら、2013
S.W. Kim, W. Suda, S. Kim, K. Oshima, S. Fukuda, H. Ohno, ..., M. Hattori
高スループットパイロシークエンシングにより明らかになったプロバイオティクス介入に対する健康成人の腸内細菌叢の頑健性
DNA Research, 20 (3) (2013), pp.241-253, 10.1093/dnares/dst006
Scopusで見るGoogle Scholar
クルンペンら、2017
E. クルンペン、N.ホフシュロア、B.ツァイス、U.ギゲンガック、E.ドーメン、R.J.ポール
活性酸素種(ROS)とミジンコの熱ストレス応答: 活性酸素を介した低酸素誘導因子1(HIF-1)と熱ショック因子1(HSF-1)の活性化とストレス遺伝子のクラスター発現
細胞の生物学, 109 (1) (2017), pp.39-64, 10.1111/boc.201600017
スコープで見るGoogle Scholar
Koteswara Raoら、2020年
K. コテスワラ・ラオ、T.V.ラクシュミ・クマール、A.クルカルニ、C.H.ホー、B.マヘンドラナート、S.デサンセッティ、...、S.サバデ
地球温暖化に対応したインドにおける暑熱ストレスとそれに伴う労働パフォーマンスの予測
Scientific Reports, 10 (1) (2020), p. 16675, 10.1038/s41598-020-73245-3
Scopusで見るGoogle Scholar
クルガノフら、2017
E. クルガノフ, 齋藤 聡, 田中 齋藤, 富永 昌彦
TRPA1の熱誘発活性化には、特定のアミノ酸への細胞外Ca(2+)結合が必要であること
生理学雑誌, 595 (8) (2017), pp.2451-2463, 10.1113/JP274083
スコープで見るGoogle Scholar
ラポルタ、2021
J. ラポルタ
ADSA財団奨学生賞: 早期からの高体温への曝露: 生産的・生理学的結果、コスト、機会
Journal of Dairy Science, 104 (11) (2021), pp.11337-11347, 10.3168/jds.2021-20722
PDFで記事を見るScopusで記事を見るGoogle Scholar
劉ら、2022年
F. リュー、W. ザオ、H.H. レ、J.J. コットレル、M.P. グリーン、B.J. リューリー、...、A.W. ベル
レビュー 豚産業における熱ストレスの影響について、我々は何を学んだか?
アニマル, 16 (2022), 10.1016/j.animal.2021.100349
Google Scholar
三上ら, 2020
A. 三上, 荻田貴志, 生井史朗, 重森慎一郎, 佐藤敏明, 下里敏明
緑茶摂取で増加する細菌Flavonifractor plautiiの経口投与はIL-17の抑制を介してマウスの急性大腸炎からの回復を促進する。
Frontiers in Nutrition, 7 (2020), Article 610946, 10.3389/fnut.2020.610946
スコープで見るGoogle Scholar
三宅ら、2016
T. 三宅, 中村修一, 趙正明, 曽健一, 井上和彦, 沼田知之, ..., 金子修一
TRPA1の低温感受性は、プロリルヒドロキシル化阻害によって誘導される活性酸素への感作によって明らかになる
Nature Communications, 7 (2016), p. 12840, 10.1038/ncomms12840
スコープで見るGoogle Scholar
宮本ら、2022
H. 宮本英樹, 浅野史郎, 石澤和彦, 須田和彦, 宮本英樹, 辻直樹, ..., 菊池純一
木材利用昆虫幼虫の炭素・窒素代謝に関与する共生細菌の潜在的ネットワーク構造
総合環境科学, 836 (2022), 第155520号, 10.1016/j.scitotenv.2022.155520
PDFで記事を見るScopusで記事を見るGoogle Scholar
宮本ら, 2022
H. 宮本博之, 河内直樹, 黒谷明彦, 守屋慎一郎, 須田和彦, 鈴木啓介, ..., 菊池純一郎
陸上養殖下流域に生育する海草の底質共生細菌構造の計算による推定
環境研究, 219 (2022), p. 115130, 10.1016/j.envres.2022.115130
グーグル奨学生
宮本・菊池, 2023
H. 宮本、菊地
解析的データ科学的アプローチによる生態系の恒常性可塑性の評価
計算構造生物学雑誌, 21 (2023), pp.869-878, 10.1016/j.csbj.2023.01.001
PDFで記事を見るScopusで記事を見るGoogle Scholar
宮本ら, 2023
H. 宮本浩次, 重田和彦, 須田和彦, 市橋洋一, 二瓶直樹, 松浦正明, ..., 平井正夫
持続可能な有機農業のための堆肥-土壌-植物相互作用の農業生態学的構造モデル
ISME Commun, 3 (2023), p. 28, 10.1038/s43705-023-00233-9
グーグル奨学生
宮本ほか, 2013
宮本英樹, 島田英樹, 佐藤智昭, 田中亮, 大島和彦, 須田和彦, ..., 松下哲也
好熱菌発酵堆肥は過酸化を防止する飼料添加物としての可能性がある
バイオサイエンス・バイオエンジニアリング, 116 (2) (2013), pp.203-208, 10.1016/j.jbiosc.2013.01.024
PDFを表示記事を表示Scopusで表示Google Scholar
中山, 1950
O.A.S.K.中山
有胞子性乳酸菌に関する研究:第1報
日本農芸化学会誌, 23 (1950), pp.
CrossRefView in ScopusGoogle Scholar
荻田ら、2020
T. 荻田, 山本恭子, 三上敦子, 重森慎一郎, 佐藤隆志, 下里敏明
フラボニフラクトールプラウティイの経口投与はマウスのTh2免疫応答を強く抑制する
免疫学のフロンティア, 11 (2020), p. 379, 10.3389/fimmu.2020.00379
スコープで見るGoogle Scholar
岡田ら、2023
S. 岡田聡, 稲生裕之, 宮本博之, 鈴木啓介, 加藤崇, 黒谷明, ..., 高橋博之
子牛の抗生物質異常症のサイレント表現型の推定
Scientific Reports, 13 (2023), p. 6359, 10.1038/s41598-023-33444-0
スコープで見るGoogle Scholar
パーデューら、1985年
S.L. Pardue、J.P. Thaxton、J. Brake
高環境温度に暴露されたヒナにおけるアスコルビン酸の役割
Journal of Applied Physiology, 58 (5) (1985), pp.1511-1516, 10.1152/jappl.1985.58.5.1511
スコープで見るGoogle Scholar
ピライら、2019
N.R. Pillai, B.M. Stroup, A. Poliner, L. Rossetti, B. Rawls, B.J. Shayota, ..., L.C. Burrage
プロピオン酸血症およびメチルマロン酸血症における肝移植: 単一施設における研究と文献レビュー
分子遺伝学と代謝、128(4)(2019)、431-443頁、10.1016/j.ymgme.2019.11.001
PDFで記事を見るScopusで記事を見るGoogle Scholar
リングゼイスとエダー、2022
R. リングゼイス、K.エダー
豚とブロイラーにおける熱ストレス: 腸-肝臓軸の障害における腸内細菌異常症の役割と、プロバイオティクス、プレバイオティクス、シンバイオティクスによるこれらの効果の回復
Journal of Animal Science and Biotechnology, 13 (1) (2022), p. 126, 10.1186/s40104-022-00783-3
スコープで見るGoogle Scholar
ローゼンバーグら, 1968
L.E. Rosenberg、A. Lilljeqvist、Y.E. Hsia
メチルマロン酸尿症: 代謝ブロックの局在とビタミンB12依存性
Science, 162 (3855) (1968), pp.805-807, 10.1126/science.162.3855.805
スコープで見るGoogle Scholar
Rosseel, 2012
Y. ロシール
lavaan: 構造方程式のためのRパッケージ
Journal of Statistical Software, 48 (2) (2012), pp.1-36, 10.18637/jss.v048.i02
Google Scholar
Rosseel et al.
Y. Rosseel、T.D.Jorgensen、N.Rockwood、D.Oberski、J.Byrnes、L.Vanbrabant、...、H.Du
潜在変数解析
Rパッケージ (2021)
バージョン0.6-9
Google Scholar
サレム他, 2022
H.M. Salem, A.H. Alqhtani, A.A. Swelum, A.O. Babalghith, S.J. Melebary, S.M. Soliman, ..., M.E. Abd El-Hack
家禽における熱ストレスとその改善におけるプロバイオティクスの役割: 最新のレビュー
Journal of Thermal Biology, 108 (2022), Article 103302, 10.1016/j.jtherbio.2022.103302
PDFで記事を見るScopusで記事を見るGoogle Scholar
佐々木ら, 2020
K. 佐々木大輔, 井上淳, 星直樹, 前田高志, 山田玲子, 近藤明彦
Bacillus coagulans SANK 70258はin vitroで潰瘍性大腸炎の微生物叢の腸内細菌を抑制し、健常な微生物叢の酪酸生成を促進する
応用微生物学とバイオテクノロジー, 104 (9) (2020), pp.3859-3867, 10.1007/s00253-020-10506-1
スコープで見るGoogle Scholar
佐藤ら、2021
K. 佐藤慶一郎, 山崎健一郎, 加藤俊一郎, 中西洋一郎, 津野哲也, 横地武内, ..., 山崎健一郎
肥満関連腸内細菌叢は尿酸の上昇を介して実験的歯周炎における歯槽骨破壊を促進する
MBio, 12 (3) (2021), p. e0077121
Google Scholar
シュナイダーズら、2020年
G.H.シュナイダーズ、J.C.フーツ、M.C.ミルフォート、A.F.A.ガリーブ、A.L.フラー、R.レカヤ、...、S.E.アグリー
熱ストレスは食肉用ニワトリの性発達を低下させ、Eimeria maximaの病原性に影響する。
Scientific Reports, 10 (1) (2020), p. 10736, 10.1038/s41598-020-67330-w
スコープで見るGoogle Scholar
清水ら, 2006
S. 清水、P.O.ホイヤー、A.ハイバリネン、A.ケルミネン
原因発見のための線形非ガウス非環状モデル
機械学習研究 (2006), pp.
スコープで見るGoogle Scholar
シディキら、2020
S.H. Siddiqui、D. Kang、J. Park、M. Khan、K. Shim
慢性熱ストレスはブロイラー小腸における熱ショックタンパク質と免疫の関係を制御する
Scientific Reports, 10 (1) (2020), p. 18872, 10.1038/s41598-020-75885-x
スコープで見るGoogle Scholar
ソンら、2014
J. Song、K. Xiao、Y.L. Ke、L.F. Jiao、C.H. Hu、Q.Y. Diao、...、X.T. Zou
熱ストレスに曝されたブロイラーの腸内細菌叢、形態およびバリア保全性に及ぼすプロバイオティクス混合物の影響
Poultry Science, 93 (3) (2014), pp.581-588, 10.3382/ps.2013-03455
PDFで記事を見るScopusで記事を見るGoogle Scholar
ストッカーら、2013
Stocker, T. F., Qin, D., Plattner, G., K., Tignor, M. M. B., Allen, S. K., Boschung, J., ... & Midgley, P. M. (2013). IPCC-2013:気候変動2013:物理科学的基礎。気候変動に関する政府間パネル第5次評価報告書へのワーキンググループIの貢献。
グーグル・スカラー
トンダら、2018
R.M.トンダ、J.K.ルバック、B.S.ランプキンス、G.F.マティス、M.J.ポス
コクシジウム症ワクチン接種および/または混合種Eimeriaチャレンジ後のブロイラー鶏の成績および腸の健康に及ぼすタンニン酸抽出物の影響
Poultry Science, 97 (9) (2018), 3031-3042頁, 10.3382/ps/pey158
PDFで記事を見るScopusで記事を見るGoogle Scholar
VaralakshmiおよびRichardson, 1983
P. バララクシュミ、K.E.リチャードソン
ラットにおけるグリコール酸からのシュウ酸合成に対するビタミンB-6欠乏および肝切除の影響
Biochimica et Biophysica Acta, 757 (1983), pp.1-7, 10.1016/0304-4165(83)90145-9
PDFで記事を見るScopusで記事を見るGoogle Scholar
ワンら、2021年
王赫朗, 興吾大, 銭毅, 孫旭, 中鍾, 陳凯朗
ジヒドロミリセチンは、ミトコンドリア機能不全を抑制することにより、乳牛乳腺上皮細胞における熱ストレス誘発アポトーシスを抑制する。
生態毒性と環境安全性, 214 (2021), Article 112078, 10.1016/j.ecoenv.2021.112078
PDFで記事を見るScopusで記事を見るGoogle Scholar
Wasti et al.
S. Wasti, N. Sah, C.N. Lee, R. Jha, B. Mishra
α-リポ酸の給与はブロイラーにおける熱ストレスの悪影響を緩和する。
PLoS One1, 16 (7) (2021), p. e0254936
CrossRefView in ScopusGoogle Scholar
渡辺ら、2018
Y. 渡辺, 福田亮介, 堤俊彦
有胞子性乳酸菌Bacillus coagulans SANK 70258の摂取が健康ボランティアの便通に及ぼす影響-無作為化二重盲検プラセボ対照並行群間比較試験-.
日本薬理治療学会雑誌, 46 (2018), pp.1549-1558
Scopusで見るGoogle Scholar
ワウタースら、2021
L. Wauters, H. Slaets, K. De Paepe, M. Ceulemans, S. Wetzels, K. Geboers, ..., T. Vanuytsel
機能性ディスペプシアの治療における有胞子性プロバイオティクスの有効性と安全性:試験的無作為化二重盲検プラセボ対照試験
Lancet Gastroenterology & Hepatology, 6 (10) (2021), 784-792頁, 10.1016/S2468-1253(21)00226-0
PDFを見る記事を見るScopusで見るGoogle Scholar
ウェスト, 2003
J.W.ウェスト
乳牛の生産に対する熱ストレスの影響
Journal of Dairy Science, 86 (2003), pp.2131-2144, 10.3168/jds.S0022-0302(03)73803-X
PDFを見る記事を見るScopusで見るGoogle Scholar
夏ら, 2009
J. Xia、N. Psychogios、N. Young、D.S. Wishart
MetaboAnalyst: メタボロームデータの解析と解釈のためのウェブサーバー
Nucleic Acids Research, 37 (Web Server issue) (2009), pp.W652-W660, 10.1093/nar/gkp356
スコープで見るGoogle Scholar
Xia and Wishart, 2011a
Xia, J., & Wishart, D. S. (2011a). MetaboAnalystを用いたメタボロームデータの処理、解析、解釈。Current Protocols in Bioinformatics, Chapter 14, Unit 14 10. https://doi.org/10.1002/0471250953.bi1410s34.
Google Scholar
Xia and Wishart, 2011b
J. Xia, D.S. Wishart
MetaboAnalystを用いたメタボロームデータからの生物学的パターン、機能、パスウェイのウェブベース推論
Nature Protocols, 6 (6) (2011), 743-760頁, 10.1038/nprot.2011.319
スコープで見るGoogle Scholar
Zhangら、2023年
X. Zhang、D. Wang、J. Liu
低酸素誘導因子-1αは泌乳乳牛の熱ストレス応答に関与する
Journal of Thermal Biology, 112 (2023), Article 103460, 10.1016/j.jtherbio.2023.103460
PDFで記事を見るScopusで記事を見るGoogle Scholarで記事を見る
引用者: (0)
© 2024 The Authors. 発行:エルゼビア社
エルゼビアのロゴとワードマーク
ScienceDirectについて
リモートアクセス
ショッピングカート
広告掲載
お問い合わせとサポート
利用規約
プライバシーポリシー
当サイトではクッキーを使用しています。クッキー設定
このサイトのすべてのコンテンツ: Copyright © 2024 Elsevier B.V., its licensors, and contributors. テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビアに帰属します。すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条項が適用されます。
RELXグループホームページ
フィードバック
この記事が気に入ったらサポートをしてみませんか?