20240319 :腱板損傷・幹細胞治療・オルソバイオロジクス

腱板損傷は一般的な上肢の筋骨格疾患であり、持続的な痛みや機能障害を引き起こす可能性があります。外科手術の臨床結果は満足のいくものであるにもかかわらず、腱板の修復には、治癒不全、癒着形成、脂肪浸潤などの問題が残されています。幹細胞は、高い増殖性、強力な傍分泌作用、および多分化の可能性を備えており、腱のリモデリングと線維軟骨の形成を促進し、生体力学的強度を高めます。さらに、幹細胞由来の細胞外小胞(EV)は、調節タンパク質やマイクロRNAを運ぶことでコラーゲン合成を増加させ、炎症や癒着形成を阻害することができます。したがって、幹細胞ベースの治療は、腱板治癒に大きな可能性を秘めた有望な治療戦略です。このレビューでは、腱板修復における幹細胞と幹細胞由来の EV の進歩を要約し、幹細胞と幹細胞由来の EV、および生体材料送達システムの根底にあるメカニズムに焦点を当てます。今後の研究では、細胞因子、遺伝子治療、新しい生体材料送達システムと組み合わせた幹細胞治療を探求する必要があります。
腱板損傷は世界中で主要な筋骨格疾患の 1 つであり、肩の痛みを訴える最も一般的な症状です ( Picavet および Schouten、2003 )。英国では、プライマリケアにおける肩の問題の有病率は 2.4% と推定されており ( Linsell et al., 2006 )、肩の痛みの 30% ~ 70% は腱板疾患に起因しています ( Mitchell et al., 2005 )。加齢、肥満、喫煙、糖尿病、遺伝学、狭い解剖学的肩峰下スペースなどの内因性因子が腱板疾患の原因となります(Titchener et al., 2014)。これらの要因のうち、加齢に伴う変性が腱板疾患の主な原因であると考えられており、一般集団では加齢に伴って腱板断裂の有病率が増加します。超音波検査による腱板全層断裂の有病率は、50代で10.7%、60代で15.2%、70代で26.5%、80代で36.6%であることが判明した(皆川ら、2013)。内因的リスクは腱板の構造的弾力性を低下させますが、職業やスポーツ活動などの外因的リスクは腱板に過度の機械的負荷を引き起こし、腱板損傷を引き起こします(Whttle and Buchbinder、2015)。腱の病理学モデルの連続体によると、機械的負荷は病理学的変化において重要な役割を果たしており ( Lewis, 2010 )、腱板腱への生体力学的負荷が繰り返されると腱板損傷のリスクが増加します ( Edmonds and Dengerink, 2014 )。 腱板損傷は、腱障害から始まり、部分的または完全な腱断裂に進行する可能性があり( Lewis, 2010 )、通常、肩の痛み、動きの喪失、および機能障害を引き起こします( Craig et al., 2017 )。一般に、腱障害は重大な問題を引き起こすことはありません。したがって、腱障害のある患者には、理学療法や鎮痛などの保存的管理コースが最初に推奨されます(Seida et al.、2010)。場合によっては、腱障害のある患者、特に高齢者の患者では腱断裂のリスクが増加する可能性があります ( Yasui et al., 2017 )。それにもかかわらず、急性肩外傷は腱の部分的または完全な断裂を引き起こす可能性があり、構造の連続性を修復するための外科的治療や、腱を骨挿入部に再付着させるための手術が必要になります。
15件の研究と腱板損傷後の371人の患者を対象とした以前の系統的レビューでは、手術を受ける時期が早ければ臨床転帰が改善することが実証された( Mukovozov et al., 2013 )。残念ながら、臨床転帰は手術後も残存しており、全体的な治癒失敗率は術後修復12か月時点で43%であり( Rashid et al., 2017 )、高齢者では最大90%に達した( Galatz et al., 2004 )。 腱板関節鏡視下修復後のリハビリテーションプロセスは通常数か月続き、アスリートはスポーツに戻るまでに 6 か月以上かかります ( Thigpen et al., 2016 )。さらに、損傷部位での瘢痕組織の形成は、組織の癒着や関節の硬さだけでなく、機械的特性の低下を引き起こす可能性があり、再断裂のリスクを高めます ( Thomopoulos et al., 2010 )。これらの問題のため、過去 10 年間、腱板の修復と再生を強化するために幹細胞を調製することへの関心が高まっています。間葉系幹細胞(MSC)は、複数の組織へのアクセス可能性、抗炎症特性、栄養因子の分泌、および再生中の組織を再細胞化するための腱細胞への分化能力により、最も人気のある幹細胞です(Lim et al., 2019)。
腱板の構造と治癒
回旋腱板は、棘上筋、棘下筋、肩甲下筋、小円筋という 4 つの筋肉で構成され、肩関節を包み込み、腱を介して上腕骨頭に密接に付着しています ( Escamilla and Andrews, 2009 ; Oliva et al., 2015 )。これらの筋肉は、肩関節の運動中の動きと動的安定化の両方において重要な役割を果たします ( Lin et al., 2018 )。腱板の特殊な解剖学的構造と血管の欠如により、簡単または効果的に治癒できない損傷が発生する可能性があります ( Hegedus et al., 2010 )。腱板損傷には、筋肉、腱、骨の複合体全体が関与することが多く、その中でも腱および腱と骨の境界面が最も頻繁に損傷し、関係する部位です。
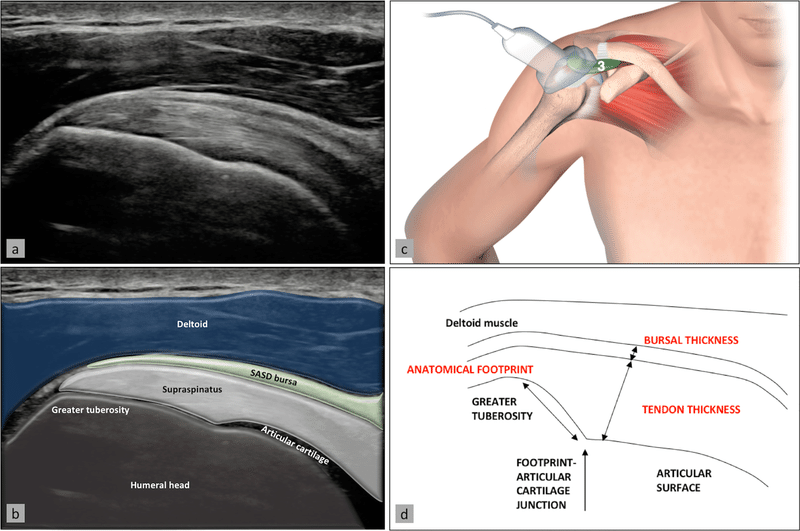
腱は、筋肉の収縮力を骨格に伝達して姿勢を維持したり、動きを生み出したりする独特の形態の結合組織です。それは常在細胞と細胞外マトリックス (ECM) で構成されます。コラーゲン線維の内側に位置する主な種類の細胞である腱細胞は、コラーゲン I 分子と ECM 分子を生成します ( Nourissat et al., 2015 )。一般に腱由来幹細胞(TDSC)とも呼ばれる腱幹/前駆細胞(TSPC)があり、分化と増殖を通じて腱細胞を再生して恒常性を維持することができます(Bi et al.、2007)。ECM には、腱特異的コラーゲン I に関与するコラーゲン、エラスチン、プロテオグリカン、糖タンパク質などの多数の分子が含まれています。三重らせん状のコラーゲン I 分子は原線維に集合し、次に線維、束を形成し、最終的には組織を形成します。 腱、このうち、コラーゲン原線維は力伝達の基礎であると考えられています ( Kannus, 2000 )。
腱と骨のユニットは付着部と呼ばれる特殊な構造であり、柔らかい腱組織と硬い骨組織の間の移行部を表します ( Yang and Temenoff, 2009 ; Andarawis-Puri et al., 2015 ) 。この組織には、骨芽細胞、骨細胞、破骨細胞、線維軟骨細胞、腱細胞などのさまざまな特殊な細胞型が常在しています。この付着部は、腱、非石灰化線維軟骨、石灰化線維軟骨、および骨の 4 つの連続的だが別個のゾーンに分割されています ( Thomopoulos et al., 2010 )。ゾーン 1 (腱領域) は、主にコラーゲン I 線維と少量のデコリンで構成されています。ゾーン 2 (石灰化されていない線維軟骨) には、主にコラーゲン II および III 線維と、少量のコラーゲン I、IX、および X コラーゲン線維が含まれています。デコリンや骨材も存在します。ゾーン 3 (石灰化線維軟骨) では、線維軟骨が石灰化され、軟骨下骨層に挿入されます。アグリカン成分とミネラル成分の両方が細胞外マトリックス組成物中に存在します。ゾーン 4 (骨領域) は、主に、骨芽細胞、骨細胞、および破骨細胞で石化されたコラーゲン I 繊維を含む骨様の組成物です。微細構造の段階的な変化により、機械的ひずみ、応力分布、効率的なエネルギー遷移が可能になります ( Rossetti et al., 2017 ; Takeshi et al., 2017 )

腱板断裂後、損傷部位は、炎症、増殖、リモデリングという 3 つの重複段階を含む自然治癒プロセスを経ます ( Docheva et al., 2015 )。炎症段階では、炎症性細胞は、好中球、単球、マクロファージなどの炎症誘発性サイトカインによって損傷部位に引き寄せられ、インターロイキン (IL)-6 や IL-1β などの炎症性サイトカインを生成します ( Lin et al.、 2004年)。さらに、組織修復を促進するために、塩基性線維芽細胞成長因子 (bFGF)、骨形成タンパク質 (BMP)、トランスフォーミング成長因子ベータ (TGF-β)、血管内皮成長因子 (VEGF) などのさまざまな成長因子が細胞から放出されます。 ( Docheva et al.、2015 )。血管新生因子は、新しく形成された線維組織への血液供給を処理する新生血管ネットワークの形成を誘導する( Fenwick et al., 2002 ; Hegedus et al., 2010 )。一方、腱細胞は損傷部位に動員され、増殖が誘導されます。第 2 段階は、ECM 成分の豊富な合成活性によって特徴付けられます。主にコラーゲンIIIであり、動員された線維芽細胞によって指示され、腱の整列の乱れと機械的弱さを引き起こします。リモデリング段階では、細胞密度と ECM 成分の合成の両方が減少します。同時に、コラーゲン III はコラーゲン I に徐々に置き換えられ、腱の ECM がより整列するように誘導されます。その間、腱の硬さと引張強度は損傷前のレベルに回復します ( Voleti et al., 2012 )。
治癒プロセスには、内因性および外因性の両方の治癒プロセスが含まれます ( Longo et al., 2011 )。外因性治癒プロセスが最初に起こり、これには周囲の鞘および滑膜からの外因性細胞の侵入が含まれ、その結果、天然付着部に代わる瘢痕組織が形成されます。瘢痕組織の形成と線維軟骨の欠如により、コラーゲン I 線維ではなくコラーゲン III 線維が分泌されます。次に、固有の治癒が活性化され、同時に腱細胞の動員、増殖、コラーゲン I 線維の分泌が誘導され、腱の機械的特性が強化されます ( Docheva et al., 2015 )。瘢痕組織にはミネラル分布の勾配がなく、コラーゲンIII線維の直径はコラーゲンI線維の直径よりも小さい( Hexter et al., 2017 )。このため、構造は弱い機械的特性を示します。コラーゲンIII線維はコラーゲンI線維に置き換えることができますが、治癒プロセスが完了するまでに通常は最大12か月かかり、再断裂の可能性が高くなる可能性があります(Lee et al., 2019 ; Haleem et al., 2021))。
腱板損傷に対する幹細胞
幹細胞ベースの治療
MSC の使用は、再生医療における有望かつ優れた治療効果です。幹細胞は、骨髄、脂肪組織、腱、臍帯血、滑液包、尿などの複数の組織から採取することに成功しています。
骨髄由来間葉系幹細胞
骨髄由来間葉幹細胞(BMSC)は、最初に発見された間葉幹細胞であり、多系統分化能力を備えた多能性細胞( Heo et al., 2016 )として機能します( Docheva et al., 2007 ; Dai et al., 2015)。;Perucca Orfei et al., 2019)脂肪細胞、骨芽細胞、軟骨細胞、および腱細胞に分解します。したがって、それらはさまざまな組織の修復および再生手順に使用されてきました。腱板損傷におけるBMSCの主な発生源は、腸骨稜および上腕骨近位部から採取できる自家細胞です。
谷口ら。(2015)は、関節鏡視下表面保持 (ASH) 修復中にフットプリントに骨髄刺激を適用すると、特に大規模な断裂において、術後の磁気共鳴画像法による Sugaya の分類に基づいてカフ修復の完全性が向上したと報告しました。症例対照研究では、BMSC強化修復と関節鏡検査中のみの修復の治癒率はそれぞれ100%と67%でした。さらに、BMSCの増加により、10年間の追跡調査後のさらなる断裂が防止されます(Hernigou et al.、2014)。
脂肪由来幹細胞
近年、脂肪由来幹細胞(ADSC)を使用して腱板修復を強化することは、その容易な取得と、微小環境および抗炎症特性を調節することによって骨形成分化を阻害する能力により魅力的となっている(Bunnell et al., 2008) ;國分ら、2020)。ADSC はアクセスしやすいため、再生療法における幹細胞の理想的な供給源です。それらは、皮下脂肪組織 ( Bunnell et al., 2008 ) および脂肪吸引の吸引物 ( De Francesco et al., 2015 )から大量に分離できます。未分化幹細胞として、それらは高い増殖率を有し、成長因子および機械的ストレスにより腱細胞に分化する可能性がある(Dai et al., 2015 ; Rinella et al., 2018)。
さらに、ADSC は腱板の再生において BMSC と同様の治療効果を示しています。急性腱板損傷と比較して、慢性腱板損傷は骨量の減少と構造的特性の低下を引き起こします。損傷部位に移植されたADSCは、上腕骨近位部の骨密度を増加させ、慢性断裂の修復における腱と骨の治癒を促進する可能性があります(Kaizawa et al., 2019 ; Rothrauff et al., 2019 ; Shin et al., 2020)。
さらに、肩甲下筋の筋腱接合部領域への ADSC の注射は、筋電図評価によって筋肉機能を改善し、筋肉の脂肪浸潤を減少させることができ、慢性腱板断裂における破損負荷を高める傾向があります ( Oh et al., 2014年)。
ADSC は、腱板損傷の臨床研究で最も一般的に使用される幹細胞の一部です。Kimらによるコホート研究。(2017) は、関節鏡視下腱板修復中にフィブリン接着剤を充填した ADSC (4.46 × 10 6細胞)の注射で治療した 182 人の患者が、再裂率の観点から評価された構造的転帰を大幅に改善できることを明らかにしました。また、MRI の結果は、再裂率が低下することを示しました。 ADSC グループの割合は、手術後少なくとも 12 か月の時点で対照グループの割合よりも低かった (それぞれ 14.3% と 28.5%)。しかし、28か月の追跡調査時点では、痛みの強さ、可動域、自己申告の機能に有意差はありませんでした。
症候性部分層腱板断裂(sPTRCT)の患者にとって、手術は第一選択の治療法ではない可能性があります。したがって、多くの研究が再生戦略として ADSC 療法を使用してきました。パイロット RCT 研究では、少なくとも 6 週間理学療法治療に反応しなかった sPTRCT 患者は、無修飾の自己脂肪由来再生細胞 (UA-ADRC) (11.4 × 10 6細胞)の単回注射を受けるように無作為に割り当てられました。 )またはメチルプレドニゾロン 80 mg(40 mg/ml; 2 ml)と 0.25% ブピバカイン 3 ml の単回注射。UA-ADRC グループの患者は、コルチコステロイド グループの患者と比較して、治療後 3 か月および 12 か月の時点で、米国肩肘外科医の標準肩評価フォーム(ASES)で有意に高い合計スコアを示しました。治療後12か月の追跡調査では、UA-ADRCの注射に関連する重篤な有害事象は報告されなかった( Hurd et al., 2020 )。遡及的比較研究では、sPTRCT患者に対するADSCの高用量(1.0×10 8細胞)の腱内注射により、治療後最長2年間、肩機能スコアと腱板強度が改善できることが示されました。MRIの結果、滑液包側の欠損は1年後にはほぼ消失し、最長2年間は再発しなかったことが示されました。重要なのは、最低2年間の追跡調査では治療に関連した有害事象はなかったということである( Jo et al., 2020 )。
腱幹/前駆細胞
腱由来幹細胞(TDSC)としても知られる前駆/腱幹細胞(TSPC)は、2007年にヒトおよびマウスから単離および同定された( Bi et al., 2007 )。TSPCは、関節鏡視下腱板修復処置中に棘上筋の腱および上腕二頭筋長頭から採取および単離できるため、そのように名付けられました(Tsai et al., 2013 ; Dei Giudici and Castricini, 2020)。他のMSCと同様に、TPSCは、高いクローン原性、自己再生能力( Al-Ani et al., 2015 )、および腱細胞、軟骨細胞、骨細胞、脂肪細胞を含む多分化能という特徴を持っています( Zhang and Wang, 2010 ; Leonardi et ) al.、2021)。彼らは、 COL1A1、テネイシンC(TNC)、強膜症(Scx)、テノモジュリン(TNMD )などの腱関連遺伝子を高度に発現しており、これらが自発的な腱性分化に寄与している可能性があります(Guo et al., 2016)。したがって、TPSC は腱再生の有望な供給源です。しかし、腱に豊富に存在するため、自己 TSPC を入手することは困難であり、臨床研究での応用が制限される可能性があります。
いくつかの研究では、前臨床研究で腱障害を治療するための TPSC の利用が調査されています ( Song et al., 2018 )。シェンら。(2012)ウサギモデルにおける腱板修復を増強するために、同種異系 TSPC を播種した足場を使用しました。誘発された免疫応答はなく、術後 12 週間の非 TSPC 治療対照修復と比較して、初期修復時のリンパ球浸潤が減少し、組織学的および生体力学的特性が改善されました。さらに、腱板由来の細胞シートを移植すると、付着部での軟骨再生と血管新生が促進され、術後4週間でVEGFおよびCOL2A1遺伝子の発現が上方制御され、術後8週間で最終失敗負荷が増加することが示された(Haradaら) al.、2017)。
臍帯由来間葉系幹細胞
臍帯由来間葉幹細胞(UCB-MSC)は、入手が容易で、増殖能力が高く、免疫原性が低いため、ヒト細胞の有望な供給源である(Wang et al., 2009 ; Bai et al., 2016)。UCB-MSCの利点は、同種異系幹細胞が骨髄穿刺や脂肪組織などの自己組織を必要としないことである( Kasper et al., 2009 )。したがって、UCB-MSCは治療前の早期に調製することができ、幹細胞の機能は患者の年齢や疾患の影響を受けません。パークら。(2015) は、急性肩甲下筋腱断裂のウサギに超音波ガイド下での UCB-MSC 注射を導入し、UCB-MSC が回旋腱板腱断裂の部分的な再生を促進し、組織学的外観、腱のサイズ、および歩行能力を改善することを明らかにしました。次に、修復のない慢性全層回旋腱板腱断裂に対するUCB-MSCの有効性を調査し、治療4週間後の組織学的検査と歩行の動作分析において、UCB-MSCの注射が同様の治療効果を示したことを発見した(Rak Kwon)ら、2020)。ただし、UCB-MSCの高用量と低用量(それぞれ2×10 6細胞と1×10 6細胞)の間に差はなく、UCB-MSCの利点が用量依存的なものではないことを示しています。 (クォンら、2019 )。別の研究では、UCB-MSCを播種した生体模倣ヒドロキシアパタイト勾配足場が、コラーゲン組織、軟骨形成、および正常な腱-骨界面と同様の生体力学的特性の改善という点で、ラット修復モデルの腱板の腱-骨界面を再生した。
嚢由来細胞
これまでの研究では、肩峰下滑液包が腱治癒のための多能性幹細胞の効力の重要な供給源であることが示唆されている(Utsutani et al., 2013 ; Baldino et al., 2020)。滑液包由来幹細胞(B-MSC)は、ルーチンの腱板修復手術から採取できる、容易に入手できる幹細胞です(Baldino et al., 2020)。他の幹細胞と同様に、B-MSC は、インビトロで高い増殖能力と分化可能性を示します( Utsutani et al., 2013 )。
別の研究では、ヒト滑液包から単離されたB-MSCは、 in vitroおよびin vivoで骨芽細胞系、脂肪生成系、軟骨形成系、腱形成系などの多系統分化によって特徴付けられることが判明した。さらに、それらは、異なる環境または前処理の下で、骨、線維軟骨、および腱に誘導されました(Song et al.、2014)。B-MSCと比較して、滑液包から単離された細胞は、腱組織において優れた生着および生存を示し、細胞を受け入れなかった組織と比較して治癒組織の厚さを増加させた( Dyrna et al., 2018 )。したがって、B-MSC は腱板損傷において強力に有望な細胞であることが示唆されています。
細胞外小胞に基づく治療
MSC は、多分化能の分化能力と直接的な細胞間相互作用に加えて、パラクリン的な方法で治療機能を媒介すると考えられています。腱板損傷モデルの例では、ヒト骨髄由来幹細胞の馴化培地(CM)は、手術後の組織学的スコア、骨密度、生体力学的な張力を増加させることにより腱板の治癒を促進します(Reiner et al.、2017 ; He et al.、2021)。治療上有効な成分を探索するために、Bruno らは超遠心分離によって MSC の CM を分画することに成功し、治療用の小胞構造を発見しました ( Bruno et al., 2009 )。細胞によって自然に放出される、脂質二重層を持つこれらのナノサイズの粒子を総称して、細胞外小胞(EV)と呼びます(Théry et al., 2018)。それらは、細胞の起源に応じて、エキソソーム、微小胞、およびアポトーシス小体に分類できます。エキソソームは最小の小胞タイプ (40 ~ 120 nm) であり、後期多小胞エンドソーム (MVE) の内向き出芽に由来し、MVE と原形質膜の融合時に放出されます。EV のメカニズムは完全には理解されていませんが、幹細胞によって分泌される EV は、細胞の増殖を誘導し、血管新生を促進し、炎症プロセスを調節し、細胞のアポトーシスに影響を与えることにより、組織の修復と再生を促進すると考えられています ( He et al., 2021年)。
BMSC 由来 EV (BMSC-EV) は、筋骨格再生分野で広く使用されています。最近、ある研究では、腱板修復を促進するためにBMSC-EVのみを静脈に注射しました(Huang et al.、2020)。その結果、BMSC-EVは、腱と骨の境界面周囲の血管新生、組織学的外観、生体力学的強度を促進し、ラット腱板再建において炎症促進因子の分泌を阻害することが示されました。彼らは、BMSC-EVが治癒過程を達成するメカニズムは、血管新生シグナル伝達経路を調節し、M1マクロファージの極性化を阻害することによる、ヒト臍帯静脈内皮細胞(HUVEC)の増殖、遊走、および血管新生管形成による可能性があると考えた。 BMSC-EVの局所送達は、内因性TPSCの増殖、遊走、および腱形成分化を促進することにより、腱の再生を促進することができる(Shi et al., 2019 ; Yu et al., 2020)。
ADSC由来EV(ADSC-EV)には、再生能力と免疫調節能力があります
Fu ら(2021)は、ADSC-EV を充填したヒドロゲルを注入した腱板が、整列したコラーゲン線維と筋束を示し、機械的特性が向上したことを報告しました。さまざまなメカニズムが腱板修復における ADSC-EV に寄与している可能性があります。
多くの研究が、腱修復におけるヒト臍帯間葉系幹細胞由来の EV (HUMSC-EV) の可能性を報告しています。キムら(2022)は、注入可能なコラーゲンを含む HUMSC-EV が骨と腱の境界面でのコラーゲンの成熟を介して骨と腱の治癒を効果的に促進し、腱板修復後 4 週間で回旋筋の脂肪変性を防止できることを実証しました。ラットアキレス腱損傷モデルでは、HUMSC-EV による治療により、組織構造が改善され、腱特異的マトリックス成分が強化され、アキレス腱の生体力学特性が最適化されました。
生体材料
生体材料分野の革新は、再生医療と組織工学の発展を推進してきました。生体材料は生体吸収性であり、徐々に分解されるため、組織には再生のための十分なスペースがあり、免疫原性や副作用は局所的および組織的に無視できます ( Garg et al., 2012 )。腱板修復において、幹細胞またはEVの送達に使用される生体材料は、移植可能な送達システムと注射可能な送達システムの2つのカテゴリーに分類できます( Chen et al., 2019 ; Liu et al., 2020 )。一般に、移植可能な送達システムは組織工学足場生体材料で構成されており、細胞の付着、成長、増殖を可能にする三次元構造を持っている必要があります。さらに、注射可能な送達システムに使用される生体材料は、薬物の拡散を最小限に抑えながら、生物活性因子と細胞を標的損傷部位に運ぶと考えられています。注射による送達には侵襲性が最小限であるという利点がありますが、細胞や障害のある組織に十分なサポートを提供することはできません。
生体材料と幹細胞間の相互作用
生体材料の地形的および機械的特性は、繊維の直径、孔径、配列、表面粗さ、マトリックスの剛性など、幹細胞の増殖と腱性分化に影響を与えます。腱の機械的微小環境により、マトリックスの剛性は腱修復中に幹細胞に影響を与えます。
腱板修復に関しては、足場は送達された細胞に機械的刺激を伝達するための効果的なツールです。したがって、細胞送達においては、生体材料によって提供される機械的環境を考慮する必要があります。幹細胞の機械的刺激は腱組織の修復に不可欠であり、幹細胞の分化と増殖に影響を与えることが示されています(Wang H.-N. et al., 2020)。伸長の大きさによって、細胞の運命が異なる可能性があります。研究では、4% のストレッチにより TPSC の腱細胞への分化が促進され、COL1A1の遺伝子発現が増加することが示されました。しかし、8%ストレッチでは、 in vitroでのPPARγ、COL2A1、Sox9、Runx2などの遺伝子発現レベルの上昇から明らかなように、腱細胞への分化とは別に、TPSCの脂肪細胞、軟骨細胞、骨細胞などの非腱細胞への分化が促進された。( Wang H.-N. 他、2020 )。
さらに、BMSCの機械的刺激は、腱誘発性遺伝子および抗炎症性サイトカインの発現を有意に増加させた( Ciardulli et al., 2020 )。
腱板損傷は持続的な症状を引き起こし、運動能力と生活の質を大きく低下させます。現在、回旋筋損傷を治療するための外科手術や従来の治療法の臨床選択肢は満足のいくものではありません。腱板の特殊な構造により、治癒過程の遅延、新しく形成された組織の生体力学的強度の低下、瘢痕癒着などのいくつかの臨床問題はまだ解決されていません。したがって、幹細胞療法は、損傷した組織を修復する体の自己潜在力を活性化するため、魅力的です。幹細胞治療における主要なエフェクターとして、EV は炎症と脂肪浸潤を軽減し、細胞増殖と腱性分化を刺激し、恒常性を維持することによって治癒プロセスを促進できます。幹細胞療法は腱板修復のための有望な戦略です。
生体材料のイノベーションは急速に進化しています。したがって、安全で有効なキャリアの翻訳は、幹細胞療法の臨床応用を前進させる鍵となります。
この記事が気に入ったらサポートをしてみませんか?