抗生物質による腸内細菌叢の乱れがヒトのワクチンに対する免疫を変化させる

メインコンテンツへスキップ
セル
ログイン
検索...
論文|178巻6号、p1313-1328.e13、2019年09月05日
全号ダウンロード
抗生物質による腸内細菌叢の乱れがヒトのワクチンに対する免疫を変化させる
https://www.cell.com/cell/fulltext/S0092-8674(19)30898-0?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867419308980%3Fshowall%3Dtrue
トーマス・ヘーガン 14
マリオ・コルテーゼ 14
ナディーン・ルーファエル 14
スレンダー・クラーナ
ハナ・ゴールディング
バリ・プレンドラン 15
すべての著者を表示
脚注を表示するオープンアーカイブDOI:https://doi.org/10.1016/j.cell.2019.08.010
PlumXメトリクス
ハイライト
微生物叢の消失は、既存の免疫力が低い被験者の抗体反応を損なう
抗生物質の投与は血液中の炎症シグネチャーの増強につながる
二次胆汁酸の喪失はAP-1/NR4Aおよびインフラマソームの活性化に関連する。
統合的解析により、マイクロバイオームが免疫に及ぼす影響の多様なメカニズムが明らかになった。
まとめ
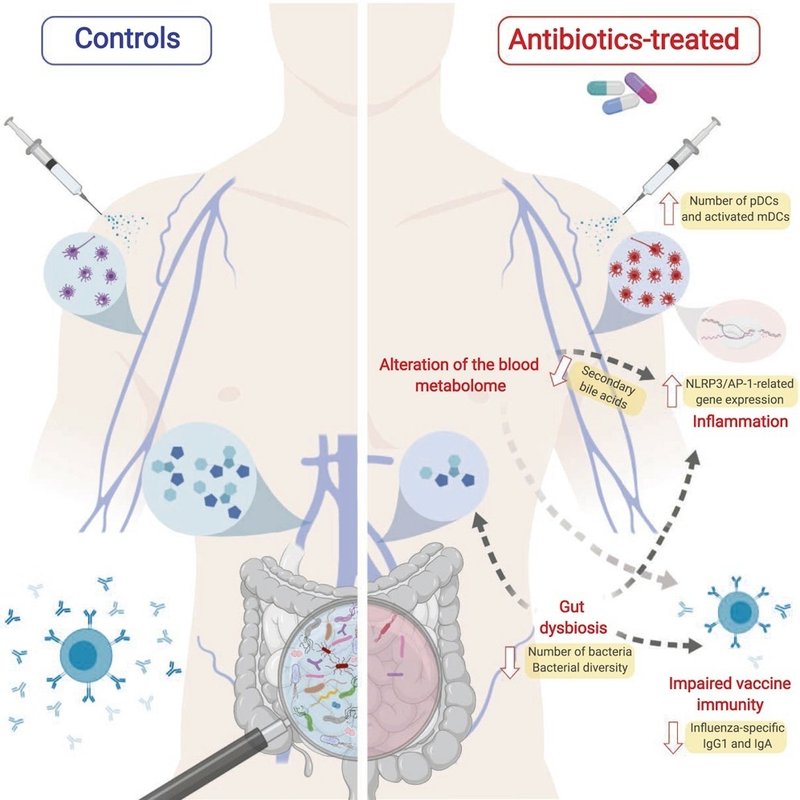
免疫においてマイクロバイオームが中心的な役割を果たしていることを示す新たな証拠が現れている。しかし、ヒトにおける因果関係の証拠は乏しい。ここでは、季節性インフルエンザワクチン接種の前後に、健康な成人に広域抗生物質を投与した。その結果、腸内細菌量は10,000倍減少し、細菌の多様性も長期にわたって減少したにもかかわらず、抗体応答には有意な影響は認められなかった。しかし、既存の抗体価が低い被験者を対象とした2つ目の試験では、H1N1特異的中和および結合IgG1、IgA反応に有意な障害がみられた。さらに、両試験において、抗生物質の投与は、(1)高齢者において以前に観察された炎症シグネチャー(AP-1/NR4A発現を含む)の増強と樹状細胞の活性化の増加、(2)血清二次胆汁酸の1,000倍減少という乖離した代謝軌跡をもたらし、これはAP-1/NR4Aシグナル伝達およびインフラムソームの活性化と高い相関があった。マルチオミクスの統合により、細菌種と代謝表現型との有意な関連が明らかになり、ヒトの免疫調節におけるマイクロバイオームの重要な役割が浮き彫りになった。
グラフィカルアブストラクト
図サムネイルfx1
大きな画像を見るダウンロード 高解像度画像
キーワード
インフルエンザ
微生物叢
ワクチン
システムワクチン学
免疫学
メタボロミクス
抗体
胆汁酸
遺伝子発現プロファイリング
システム生物学
はじめに
私たちの腸内には何兆個もの細菌が生息しており、その数は体内の真核細胞を10対1で上回っている(Savage, 1977)。宿主の代謝(Nicholsonら、2012年、Sonnenburg and Bäckhed、2016年)、腸管免疫(Belkaid and Hand、2014年、Steinhoff、2005年)、自己免疫およびアレルギー炎症(Kosticら、2014年、Mitreら、2018年、Scherら、2013年)、さらには中枢神経系とのコミュニケーション(Carabottiら、2015年)に対する多様な影響を通じて、私たちのいわゆる「第二のゲノム」であるこの微生物群集が、生理学を形成する上で強力な役割を担っていることを示唆する新たな証拠が出てきている。自己免疫やアレルギーの促進に加えて、マイクロバイオーム組成の変動は、HIV感染の予防(Klattら、2017年)や抗PD1がん免疫療法(Gopalakrishnanら、2018年、Routyら、2018年、Zitvogelら、2018年)を含む様々な免疫介入における有効性の低下と関連している。
しかし、マイクロバイオームが免疫系に与える影響を裏付ける証拠の多くは、マウスモデルでの研究やヒトでの相関研究(Belkaid and Hand, 2014, Keeney et al. マイクロバイオームがヒトの生理機能を形成するメカニズムについてより包括的な理解を深めることは、免疫疾患の治療的調節にマイクロバイオーム-宿主軸を活用する上で極めて重要である。なぜなら、多くのワクチンの有効性は地域によって大きく異なることが知られており(Collins and Belkaid, 2018, Levine, 2010, Littman, 2018)、マイクロバイオームの組成もまた地域によって大きく異なるからである(Yatsunenko et al.) したがって、腸内細菌叢がワクチン接種に対する反応性にどのような影響を与えるかを理解することは、ワクチンの有効性を高め、世界の公衆衛生を改善する上で重要な意味を持つ。
防御免疫応答を担う生物学的メカニズムの理解を深めるために、免疫学者は、システムワクチノロジーという新分野において、ワクチン接種に対する応答を包括的にプロファイリングするためのハイスループット細胞・分子技術を使用し始めている(Hagan and Pulendran, 2018, Pulendran, 2014, Pulendran et al., 2010, Rappuoli et al.) このような研究から得られた生物学的に重要な知見の1つは、季節性インフルエンザワクチンに対する免疫応答の解析において、ワクチン接種後3日目の細菌フラジェリンを認識できるToll様受容体であるTLR5の発現と、その後の28日目の抗体応答の大きさとの関連が同定されたことである(Nakaya et al.) この知見により、抗生物質投与マウスとTLR5ノックアウトマウスを用いた追跡実験が行われ、TLR5を介した腸内細菌叢からのフラジェリン感知が阻害されると、不活化インフルエンザワクチン接種に対する形質細胞応答と抗体応答が障害されることが示された(Oh et al.) このことから、腸内細菌叢がワクチンに対する体液性応答に影響を及ぼす潜在的な役割が確立された。
そこで、抗生物質投与による腸内細菌叢の減少が、ヒトのワクチン接種に対する反応に影響を及ぼすかどうかを理解するために、2シーズンにわたり3価不活化インフルエンザワクチン(TIV)を接種した抗生物質投与群と対照群において、自然免疫反応と適応免疫反応の広範なプロファイリングを行った。その結果、抗生物質投与によって腸内細菌叢が大きく変化した結果、インフルエンザに対する既存の免疫力が低い被験者において、H1N1特異的免疫グロブリンG1(IgG1)反応に有意な影響が認められた。さらに、抗生物質の投与は血漿中のメタボロームにも顕著な影響を及ぼし、微生物に関連した胆汁酸代謝の乱れは、炎症の細胞および転写シグネチャーの上昇と高い相関関係を示した。これらの知見を総合すると、ヒトの免疫調節におけるマイクロバイオームの未知の役割が明らかになった。
研究結果
抗生物質の使用は腸内細菌量の大幅な減少と細菌多様性の持続的変化をもたらす
季節性インフルエンザワクチンに対する免疫応答の形成におけるヒト腸内細菌叢の役割を調べるため、2014-2015年シーズンに18-45歳の健康な22人を登録した。11人の被験者を、ネオマイシン、バンコマイシン、メトロニダゾールの経口カクテルからなる5日間の広域抗生物質レジメンに無作為に割り付け、消化管内のグラム陰性菌(ネオマイシン)、グラム陽性菌(ネオマイシン、バンコマイシン)、嫌気性菌(メトロニダゾール)を減少させることを目的とした。抗生物質の投与はワクチン接種の3日前から1日後まで行った。免疫後1年まで定期的に生物学的サンプルを採取し、免疫系の主要な側面を測定した(図1A)。
図サムネイルgr1
図1研究の概要と健康成人の腸内細菌叢に対する抗生物質使用の影響
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
図サムネイルfigs1
図S1図1に関連するフェーズ2における健康成人の腸内マイクロバイオームに対する抗生物質使用の影響
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
我々は、抗生物質の使用が腸内細菌叢に及ぼす影響を定量化することから解析を開始した。ワクチン接種当日、抗生物質を投与された被験者では、腸内細菌の相対的存在量の代用指標である便1gあたりの16S rRNAコピー数の合計が3~4log減少した(図1B)。1日目(抗生物質投与最終日)には、抗生物質投与群の総細菌量は頭打ちとなり、その後7日目から30日目にかけてベースラインレベルまで徐々に回復した。これと同様に、フラジェリンとリポ多糖(LPS)の糞便中濃度も、抗生物質投与後の早い時点で、同様の動態で有意に減少した(図1Cと1D)。予想通り、対照群では16S rRNAコピー数、フラジェリン、LPS濃度の経時的な有意な変化は認められなかった(図1B-1D)。
総細菌量が著しく減少したことから、どの細菌種が最も影響を受けたかを検討した。ファミリーレベルの解析から、微生物群集組成の顕著な変化が明らかになり、相対的な存在量は抗生物質使用後1ヵ月まで有意に変化した。0日目、1日目、3日目では、腸内細菌科が治療対象者の腸内細菌叢を支配していたが、このコホートでは2人が妨害に抵抗しているように見え、細菌叢全体の数や組成に明らかな変化の兆候を示さなかった(図1E)。7日目まで、腸内細菌科細菌は、割合は低いものの、数人の被験者で高いままであった。Streptococcaceae(連鎖球菌科)も初期の時点ではより豊富であったが、Lachnospiraceaceae(ラクノスピラ科)、Ruminococcaceae(ルミノコッカス科)、Bacteroidaceae(バクテロイデス科)、Veillonellaceae(ベヨネラ科)は0日目から3日目にかけて減少した。30日目までに、Lachnospiraceae、Enterobacteriaceae、Ruminococcaceaeの相対レベルは乱れたままであったが、最終的には90日目までにベースラインに戻った(図1E;表S1)。
また、β(サンプル間)多様性の主座標分析(PCoA)により、全体的な微生物群集構造の違いを定量化した(図1F)。0日目(ワクチン投与日)には、抗生物質投与群のサンプルは対照群から乖離しており、3日目に両群間の距離が最も大きくなった。興味深いことに、180日目でも両群の微生物組成はかなりの非類似性を示し、抗生物質投与群では元の微生物組成が部分的にしか回復していないことが示唆された。また、α(サンプル内)多様性を調べ、抗生物質投与前後の抗生物質投与群サンプルにおける微生物の豊かさと均等性を比較した。どちらの指標も抗生物質使用後に大きく損なわれ、異なる操作分類単位(OTU)の数は3日目に最も少なく記録された(図1G)。特筆すべきことに、種の豊富さと生物多様性は6ヵ月時点でも完全には回復しておらず、先行研究(Becattini et al., 2016, Dethlefsen and Relman, 2011, Modi et al., 2014)と一致して、固有の細菌種が長期間にわたって失われていることを示している。
インフルエンザワクチン接種による抗体反応に対する抗生物質の効果
次に、腸内細菌叢に対する深く持続的な影響が、TIV免疫に対する適応免疫応答にどのように影響するかを調べた。まず、微量中和(MN)力価を測定することにより、ワクチン接種者の血清抗体応答を評価した。その結果、両コホートとも、ワクチンに含まれる3種類のインフルエンザ株それぞれに対して、すべての時点で同程度のMN力価を示したことから、抗生物質による腸内生態系の撹乱は、TIV特異的抗体がin vitroで同種のワクチン株を中和する能力を有意に変化させないことが示唆された(図2A)。30日目の血清転換率(ワクチン接種前と接種後の血清の間で力価が4倍以上上昇した被験者の割合を示す)の解析でも、2群間で同等の結果が得られた(図S2A)。しかし、全体的に血清転換の割合が低いことに気づいたが、これはおそらく一部の被験者で観察された高いベースライン力価のためであろう。
図サムネイルgr2
図2抗生物質投与がTIVに対する体液性反応に及ぼす影響
キャプション
大きな画像を見るダウンロード 高解像度画像
図サムネイルfigs2
図S2図2に関連するTIVに対する体液性応答の追加測定値
フルキャプションを表示
大きい画像を見るダウンロード 高解像度画像
抗体応答への影響の欠如が、インフルエンザに対する既存の免疫によるものであるかどうかを判定するために、1年後に、ワクチン接種前のMN力価が2015-2016年のTIV製剤に含まれる3つのワクチン株それぞれについて≦320であり、過去3シーズンのインフルエンザでTIVを受けていない11人の被験者を追加登録した。このうち 5 例を,第 1 相と同じ抗生物質レジメンを投与する群に無作為に割り付けた.本研究の2段階の参加者のベースライン時のMN力価の比較を表S2に示す。重要なことに、第2相の抗生物質投与群の5人の腸内細菌叢は、第1相と同様に微生物の豊富さと多様性が減少していた(図S1A-S1C)。しかし、この2回目の実験では、ワクチン接種後90日目と180日目に、抗生物質投与群の被験者でH1N1 A/California特異的中和能の低下も認められたが、他の2つのインフルエンザ株では差は認められなかった(図2B)。予想通り、ベースライン力価が低かったことから、2番目のコホートではセロコンバージョン率が非常に高く、3つのワクチン株のうち2つについて、対照群と抗生物質群の両方で100%に達した(図S2B)。
驚くべきことに、第2相の抗生物質投与群では、ELISA法で測定した7日目のH1N1 A/California株(図2C、右パネル)に特異的なIgG1抗体濃度が有意に低下していた。これは、IgG1反応の大きさが抗生物質処理によって強い影響を受けなかったフェーズ1とは対照的であった(図2C、左パネル)。これらの結果を確認するため、ハイスループットのLuminexベースのアッセイ(Brown et al.) 2つの独立した測定値の間には顕著な相関があり、第2相で抗生物質を投与された被験者は、Luminexアッセイによってワクチン接種後7日目と30日目にH1N1特異的IgG1力価の産生に有意な障害を示した(図2Dと2E)。ELISAデータと一致して、ルミネックスによる第1期参加者では、IgG1産生に対する抗生物質投与の有意な影響は検出されなかった(データは示さず)。ワクチン誘発性のIgG1産生は、インフルエンザに特異的なIgG応答全体の大部分を占めるが(Frasca et al.、2013)、他のIgGサブクラスの濃度が抗生物質処理によって影響を受けるかどうかを検討したが、IgG2(図S2C)またはIgG3抗体には有意な影響は認められなかった(データは示さず)。重要なことは、H1N1 A/California株に対する抗体親和性も抗生物質使用の結果として変化したことである(図2F)。
ワクチンで誘導された抗体反応の大きさに対する抗生物質処理の顕著な影響は、IgG1だけに限られたものではなかった。表面プラズモン共鳴(SPR)を用いると、第2相抗生物質投与群では、ワクチン接種前後でH1N1特異的IgA抗体応答が低いことが検出された(図2G)。これらの差はルミネックスでも確認され(図2Hと2I)、第1相抗生物質群では観察されなかった(図S2D)。
次に、適応免疫の他の重要な側面にも影響があるかどうかを検討した。季節性インフルエンザワクチン接種は、抗原特異的B細胞を形質芽細胞(PB)(Wrammertら、2008)と記憶B細胞経路にコミットする「活性化B細胞」(ABC)に分化させる(Ellebedyら、2016)。血液中のワクチン特異的PBとABCの頻度を評価したところ、対照群と抗生物質投与群の間に差は認められなかった(図S3A-S3C)。さらに、B細胞の補助に重要な血中濾胞性T細胞(Tfh)様細胞(Crotty, 2014)の頻度を評価したところ、差は認められなかった(図S3D)。さらに、ワクチン接種7日後に単一ワクチン誘発PBを選別し、本研究の両フェーズで抗生物質を投与した被験者からTIV反応性ヒトモノクローナル抗体(hmAbs)を作製することにより、B細胞体細胞超変異(SHM)に対する抗生物質使用の影響の可能性を検討した。次に、軽鎖および重鎖の変異数を、対照被験者または2014-2015インフルエンザシーズンに4価インフルエンザワクチン(QIV)を接種した健常対照者から分離したインフルエンザ陽性hmAbsの変異数と比較したが、有意差は認められなかった(図S3E)。これらの結果を総合すると、腸内細菌群集の乱れは、体液性免疫が有意に存在しない場合、インフルエンザワクチン接種に対する抗体応答を損なう可能性があることが示された。
サムネイル図3
図S3図2に関連するTIVに対する適応的細胞応答
キャプション
大きな画像を見るダウンロード 高解像度画像
ワクチン誘発血中転写シグネチャーに対する抗生物質使用の影響
ワクチン誘発転写応答の大きさと動態に対する抗生物質使用の潜在的影響を評価するために、各コホートにおいてワクチン接種後1-7日目に、0日目と比較して発現が異なる遺伝子を同定した(図3A)。反応動態は両群で同じであった:ワクチンによる遺伝子発現の変化は1日目にピークに達し、その後3日目までに減少し、7日目に2回目の増加がみられた。対照群では、抗生物質投与群に比べ、すべての時点において差次的発現遺伝子の増加が緩やかであったが、これは被験者の数がわずかに多かった(17人対15人)ためと考えられる。
図サムネイルgr3
図3対照群と抗生物質投与群におけるTIVに対する転写反応
キャプション
大きな画像を見るダウンロード 高解像度画像
次に、遺伝子セット濃縮解析(GSEA)(Subramanian et al. このために、われわれのグループが以前、一般に公開されているヒト血液トランスクリプトームデータセットの大規模ネットワーク統合によって同定した血液転写モジュール(BTM)のセットを利用した(Li et al.) 対照群と抗生物質投与群の両者とも、非常に類似した濃縮パターンを示した。これらのパターンは第1相と第2相の間でも維持されており、抗生物質投与は、既存の力価が低い被験者であっても、ワクチン接種によって誘導される転写経路を強く変化させないことが示唆された(図S4A)。1日目には、樹状細胞の活性化、抗原提示、インターフェロンシグナル伝達など、自然免疫応答に関連するモジュールの濃縮が観察された。7日目には、PBの増殖を反映する細胞周期と形質細胞関連モジュールが濃縮され、適応反応の特徴が現れた。これらの観察結果は、以前のインフルエンザシーズンにおける健常成人ワクチン接種者の解析で同定された活性化経路と一致していた(Nakaya et al.) さらに、これらの正規のインフルエンザワクチン応答経路に関与する個々の遺伝子の時間的プロファイルを調べたところ、インターフェロンシグナル伝達(図3C)や形質細胞および免疫グロブリン発現(図3D)に関連する遺伝子発現の動態や大きさに抗生物質の使用は影響しなかった。これらの所見を総合すると、抗生物質の投与はインフルエンザワクチン接種に対する転写反応にほとんど影響を与えないことが示唆される。
サムネイル図4
図S4第1相および第2相における転写および代謝反応の比較(図3、5、6に関連
キャプション
大きな画像を見るダウンロード 高解像度画像
抗生物質の使用は血液中の炎症性転写および細胞応答を誘導する
次に、抗生物質の投与だけで血液中のトランスクリプトームに変化が起こるかどうかを調べた。抗生物質を投与された被験者のスクリーニングから0日目までの遺伝子発現の変化に基づいてGSEAを行った(図4A)。その中には、炎症性シグナル伝達の増加や樹状細胞の活性化、B細胞や細胞周期に関連する発現の減少などが含まれ、これらは研究の両段階においてほぼ一貫していた(データは示さず)。また、フローサイトメトリーでいくつかの自然免疫細胞集団の頻度を測定したところ、抗生物質投与後、CD80+ CD16+骨髄性樹状細胞(mDC)とCD123++形質細胞様樹状細胞(pDC)という2つの樹状細胞サブセットの有意な増加が観察された(図4B)。これらの変化は、遺伝子発現によって検出された樹状細胞活性化の濃縮と一致していた(図4A)。両集団は抗生物質投与期間中(0日目と1日目)、抗生物質投与被験者で上昇し、抗生物質投与中止後はコントロールレベルに戻り始めたことから、これらの変化は抗生物質が誘発した腸内細菌叢の擾乱によって引き起こされたことがさらに示唆された。
図のサムネイルgr4
図4抗生物質投与に対する転写および細胞反応
キャプション
大きな画像を見るダウンロード 高解像度画像
抗生物質に反応して活性化された経路の中で特に興味深かったのは、AP-1(FOS/JUN)とNur(NR4A)転写因子に関連するいくつかのモジュールであった(図4Cと4D)。これらのモジュールは、ワクチン接種後1日目の抗生物質投与被験者では上昇したままであったが、ワクチン接種後の対照被験者では低下していた(図4C)。驚くべきことに、これらのモジュールの動態を、2010-2011年のインフルエンザシーズンにワクチン接種を受けた若年および高齢者のものと比較したところ(それぞれ水色と緑色のバー)(Nakaya et al.、2015)、若年被験者が対照被験者と同様に1日目にこれらのモジュールの発現低下を示したのに対し、抗生物質投与被験者はワクチン接種後にこれらの経路の発現上昇を示した高齢ワクチン接種者と同様の挙動を示した。これらのモジュールには、抗生物質投与後に誘導されたインターロイキン6(IL6)、IL8、CXCL2を含むいくつかの炎症性サイトカインの遺伝子も含まれていた(図4D)。これらの転写応答を制御するAP-1とNR4Aの潜在的役割をさらに調べるために、TRANSFACデータベース(Matys et al. これらの転写因子とそれぞれの標的遺伝子のいくつかとの間には、1日目に強い相関が観察された(図4E)。これらの結果は、抗生物質による腸内細菌叢の減少が、加齢に伴う免疫応答の変化と一致する形で、自然免疫細胞における炎症性シグナル伝達を促進する可能性を示している。
抗生物質の使用は血中メタボロームを乱す
腸内細菌叢は、複合糖質の消化やビタミンの合成を助けるなど、食事中の様々な栄養素の代謝において重要な役割を果たすことがよく知られている(Levyら、2017、Nicholsonら、2012、Tremaroli and Bäckhed、2012)。そこで我々は、アンターゲットの高分解能メタボロミクスにより、抗生物質使用後の被験者の血中メタボロームの変化を検出できるかどうかを検討した。その結果、抗生物質を投与した被験者のメタボロームには、対照被験者に比べて大きな変動が認められ(図5A)、最も変動が大きい代謝物の特徴について、スクリーニング時と0日目の時点間のユークリッド距離で測定したところ、対照被験者に比べて大きな変動が認められました。mummichogソフトウェア(Li et al., 2013)を用いた差分特徴のエンリッチメント解析により、胆汁酸代謝やトリプトファン代謝を含む複数の経路において、抗生物質による変化が明らかになった(図5B)。これは、胆汁酸プールの制御における腸内細菌叢の確立された機能(Ridlonら、2014)、ならびにインドール、キヌレニン、およびセロトニン経路に向かうトリプトファン代謝の制御(Agusら、2018)と一致する。我々のアンターゲットアプローチでは特定の代謝物を確実に同定することができないため、抗生物質投与後の個々の胆汁酸の変化をさらに特徴付けるために、有効な化学標準物質を用いて独立した液体クロマトグラフィー質量分析(LC-MS)分析を行った。微生物叢は肝臓で合成された一次胆汁酸を腸内で二次胆汁酸に変換する役割を担っていることから(Ridlon et al.、2006)、抗生物質投与後には一次胆汁酸と二次胆汁酸の比率が上昇すると予想された。実際、抗生物質を投与した被験者の血清中では、一次胆汁酸レベルの上昇と二次胆汁酸レベルの劇的な減少が見られた(図5C)。
図5
図5抗生物質投与とインフルエンザワクチン接種が血中メタボロームに及ぼす影響
キャプション
大きな画像を見るダウンロード 高解像度画像
次に、腸内細菌叢の減少がインフルエンザワクチン接種に対する代謝反応に及ぼす影響を調べた。対照被験者では、ワクチン接種に対する代謝の変化は1日目にピークに達した(図5D)。抗生物質を投与した被験者では、1日目の反応の大きさは同程度であったが、対照被験者を超えてメタボロームの大きなシフトが維持され、ワクチン接種後7日目でも0日目と比較してかなりの差が認められた。ワクチン接種後に濃縮された代謝経路を調べたところ、2群間で濃縮経路の重複はほとんど見られなかった(図5E)。このように、抗生物質は血液中のインフルエンザワクチン接種に対する代謝反応を有意に変化させた。
代謝反応の違いをさらに調べるために、すべての時点にわたって、どちらのグループでも異なる量の代謝物ピーク(p < 0.01)のfold-change値について主成分分析(PCA)を行った。その結果、抗生物質投与群では、ワクチン接種前と接種後の両方で、対照群と比較して代謝経路が大きく変化していた(図5F)。このような差は、研究の両段階において観察された(図S4C)。重要なことは、抗生物質を投与された2人の被験者の代謝軌跡が対照被験者と同じグループであったことであり、この2人はマイクロバイオームにほとんど変化を示さなかった(図1)。
二次胆汁酸の攪乱は、抗生物質投与被験者におけるNLRP3インフラマソームシグナル伝達の上昇と関連している。
腸内細菌叢の消失が血漿中メタボロームを強く変化させたことから、この代謝障害と抗生物質投与後に観察された細胞および転写変化との関連性を探ろうと考えた(図4)。特に、抗生物質を投与した被験者で有意に変化した胆汁酸代謝異常は、炎症性腸疾患(IBD)と関連しており(Pavlidis et al. 実際、トランスクリプトームおよびメタボロームデータの統合解析により、個々の二次胆汁酸レベルと、抗生物質投与後に誘導された図4で同定された炎症性BTMの多くとの間に有意な逆相関が示された(図6A)。抗生物質投与後に血漿中で1,000倍の減少を示したリトコール酸(LCA)(図6E)は、抗生物質投与被験者ではワクチン接種後に発現が上昇したが(図6C)、対照被験者では発現が低下した(図6D)M35.0などのAP-1シグナル伝達に関与するモジュールを含む炎症反応と最も強い関連を示した。LCAは胆汁酸受容体TGR5の最も強力なアゴニストであり(Kawamataら、2003)、TGR5シグナル伝達はマウスにおいてNLRP3インフラムソームの活性化を阻害することが示されている(Guoら、2016)。さらに、AP-1はIL-1Bを介したIL-6などの炎症性サイトカインの誘導に重要な役割を果たすことが知られている(Cahill and Rogers, 2008)。このように、我々はまた、インフラマソームおよびIL-1Bシグナル伝達に関連する遺伝子の発現変化を調べたところ、LCAおよび他の二次胆汁酸との強固な関連を見出した(図6Aおよび6B)。これらの知見は、マイクロバイオームがヒトの炎症反応を制御する可能性のあるメカニズムを浮き彫りにした。
図のサムネイルgr6
図6二次胆汁酸の攪乱は、抗生物質を投与した被験者におけるNLRP3インフラマソームシグナル伝達の上昇と関連している。
キャプションを表示する
大きな画像を見るダウンロード 高解像度画像
MMRN解析は、腸内マイクロバイオームが異なる経路を通じて炎症シグナル伝達とワクチン接種に対する抗体反応を制御していることを示唆している。
腸内細菌叢は、微生物刺激が全身に移行することによって直接的に、あるいは代謝産物などの二次メッセンジャーを介して間接的に、免疫応答に影響を与える可能性がある(Brenchley et al. そこで、血清中のフラジェリンと抗LPS抗体の濃度を測定したが(図S1D-S1F)、対照群と抗生物質投与群との間に差は認められず、微生物産物の末梢への移行は否定された。推定される間接的影響を探索するために、我々は、抗生物質使用中およびインフルエンザワクチン接種中のこれらのデータタイプ間の関連を同定するために、転写、代謝、マイクロバイオーム、および細胞測定を統合した、以前に記載されたように(Liら、2017;STAR Methodsおよび図S5も参照)、マルチスケール、多因子応答ネットワーク(MMRN)を構築した。細菌-代謝物間の関連は、ネットワーク内の最も有意な関連を支配しており(図7A)、腸内細菌叢が宿主の代謝を形成する実質的な能力を示している。
サムネイル図5
図S5 図7に関連するMMRN構築のワークフロー
キャプション
大きな画像を見るダウンロード 高解像度画像
サムネイルgr7
図7MMRN解析から、炎症シグナル伝達およびH1N1特異的IgG1応答の制御における腸内細菌叢の異なる機能が示唆される
キャプションを表示
大きな画像を見るダウンロード 高解像度画像
次に、観察された胆汁酸代謝およびH1N1特異的IgG1応答の違いに最も関連するネットワークノードを同定するために、我々は、抗生物質によるLCAの変化またはH1N1特異的IgG1の30日目の存在量のいずれかと、その0日目対スクリーニングの倍数変化の相関によってランク付けされた特徴を用いて、濃縮ベースのアプローチでネットワークを照会した(図7B)。前回の解析と一致して、LCAの減少と、炎症反応、樹状細胞活性化、AP-1シグナル伝達に関与する多くのBTMを含む転写クラスターG3の増加との間には、強い負の相関が見られた。さらに、LCAは、胆汁酸生合成に濃縮された代謝物クラスターM79とも密接に関連していたことから、我々のアンターゲットメタボロミクス解析は、マイクロバイオームの摂動によって変化した代謝経路を正確に捉えていることが示唆された。
抗生物質を投与した被験者では、胆汁酸代謝とIgG1反応の両方が障害されているにもかかわらず、LCAとH1N1特異的IgG1量の両方に関連する代謝クラスターと転写クラスターの重複はほとんど見られなかった。アンドロゲンおよびエストロゲン代謝に富む唯一の代謝物クラスターM84が存在し、これは両パラメーターと負の相関を示した。この低い重複度と一致して、IgG1力価は胆汁酸の存在量の変化と弱い相関を示した(データは示さず)。代わりに、共通に関連する特徴は細菌に支配されており、3つの細菌クラスターが両パラメーターと正の相関を示した。これらのクラスターの組成分析では、Lachnospiraceae科とRuminococcaceae科に多く含まれることが示され(図7C)、両科とも抗生物質投与後に激減した(図1E)。興味深いことに、IgG1応答と特異的に関連するクラスターの中には、免疫機能の重要な調節因子として浮上している脂肪酸代謝に高度に濃縮された2つのクラスター(M26とM58)があった(Ganeshan and Chawla, 2014, Sinclair et al.)
ネットワークにおける細菌クラスターの中心的な役割から、腸内細菌量がこれら2つの経路に及ぼす重要性をさらに検討することになった。実際、LCAの変化は、便中の細菌量、フラジェリン、およびLPS含有量と強く関連しており(図7D)、H1N1特異的IgG1の存在量も、抗生物質投与後のフラジェリンの減少と有意に相関していた(図7E)。興味深いことに、フラジェリンとIgG1力価の間のこの正の相関は、ベースライン力価が低いフェーズ2の被験者においてのみ有意であり、重篤なディスバイオシスに対する免疫記憶の回復力を浮き彫りにした。これらの結果から、二次的な胆汁酸関連炎症シグナル伝達とインフルエンザワクチン接種に対するIgG1応答の障害は、抗生物質による腸内主要細菌群集の消失によって媒介される独立した効果として生じることが示唆される。
考察
ベースラインの中和抗体レベルが異なる2つのコホートで本試験を実施することにより、既存の体液性免疫がワクチン接種に対する適応反応の微生物学的媒介による変化に及ぼす影響について洞察することができた。ベースライン抗体価の高い被験者では、抗体反応への影響は最小限であったが、ベースライン抗体価の低い被験者の2番目のコホートでは、ワクチン接種後のH1N1特異的IgG1の量に対する抗生物質投与の顕著な影響が観察された。注目すべきことに、この効果はH1N1に対する反応でのみ観察され、他の株に対しては観察されなかった。この理由は今のところ不明であるが、成人におけるワクチン接種後のH3N2およびB抗体反応は、主に想起記憶反応である可能性がある。最近の研究では、ワクチン接種または感染による事前の曝露により、成人はH3N2およびB亜型の記憶が高いことが示されている(Khurana et al.)
これらの結果は、TLR5ノックアウトまたは抗生物質投与によりTLR5を介したフラジェリンの感知が失われると、インフルエンザワクチン接種後の抗体価が有意に低下するという、マウスを用いた我々の以前の知見(Oh et al. これらの研究で使用されたマウスはインフルエンザに対してナイーブであったが、事実上すべてのヒトは過去にインフルエンザに罹患した経験があり、一次的な反応と比較して、想起反応に関与するメカニズムは腸内細菌叢の変化に対してより強い可能性がある。これらの結果は、ヒト免疫系の真の適応的性質を浮き彫りにしている。ヒト免疫系は、微生物叢の摂動に起因する著しい生理的障害にもかかわらず、刺激やチャレンジに対する適切な応答をほぼ維持することができるようである。これはまた、免疫系の形成における免疫記憶と環境の重要性を浮き彫りにしている。これまでの研究で、ヒト集団内の免疫の変動は、遺伝的要因よりもむしろ、過去の免疫曝露などの環境要因によって主に引き起こされることが示されている(Brodin et al.) 本研究の結果は、成人の免疫応答は免疫歴によってほぼ決定され、マイクロバイオームの一過性の変化に強いという概念と一致している。しかし、このような解析を、免疫系が未成熟で刷り込みの影響を受けにくい乳幼児集団にまで拡大することは、今後極めて重要になるだろう。このことは、喘息や1型糖尿病などの自己免疫疾患と、乳児における分娩方法に関連した腸内微生物の違いとの相関関係についての報告によって、すでに明らかになっている(Dominguez-Belloら、2010、Giongoら、2011、Thavagnanamら、2008)。
自然免疫応答に関しては、抗生物質の投与のみで、免疫系内の細胞および転写に大きな変化が生じ、炎症性状態が促進されることが観察された。統合的解析により、これらの炎症反応は、細菌を介した二次胆汁酸の産生低下と強く関連していることが明らかになった。TGR5が介在するNLRP3インフラマソームの阻害(Guoら、2016年)に加え、LCAなどの二次胆汁酸は、ビタミンD受容体(Bakke and Sun、2018年、Makishimaら、2002年)およびプレグナンX受容体(Shahら、2007年、Staudingerら、2001年)の両方に結合することにより、炎症性サイトカイン産生および核因子κB(NF-κB)標的発現を抑制することも示されている。これらの結果を総合すると、抗生物質投与後に末梢血で検出される炎症反応は、腸内細菌叢による胆汁酸代謝の障害の結果として、インフラマソームシグナル伝達の亢進によって引き起こされることが示唆される。
興味深いことに、抗生物質投与によって誘導される炎症性状態は、高齢者におけるインフルエンザワクチン接種に対する初期転写反応と類似性を示した。マイクロバイオームの構成は加齢とともに変化することが知られており、ファーミキューテス門と全体的な多様性が減少している(Salazar et al.) 微生物叢の加齢に伴う変化が胆汁酸代謝に及ぼす影響についてはよくわかっていないが(Frommherz et al.、2016)、高齢者の腸内細菌叢は短鎖脂肪酸などの他の免疫調節代謝産物を産生する能力が低いという証拠がある(Rampelli et al.、2013)。これらのアップレギュレートされた経路(AP-1およびNR4Aシグナル伝達)は、細菌のLPSによっても誘発されることが知られており、腸管透過性はマウスにおいて加齢とともに増加することが示されている(Thevaranjanら、2017)。腸内細菌叢が高齢者の免疫系に影響を及ぼす直接的・間接的メカニズムについてさらに調べることは、加齢に伴うマイクロバイオームの変化が慢性炎症や免疫老化にどのように寄与するかを理解する上で価値があるだろう。
最後に、抗生物質治療とインフルエンザワクチン接種に対する反応を複数のデータ型にわたって統合したMMRNを構築した。その結果、マイクロバイオームが、抗体応答への影響とは異なるメカニズムで、胆汁酸代謝とその結果としての炎症への影響を調節していることが示唆された。興味深いことに、IgG1応答は、免疫応答の重要なオーケストレーターとして浮上している脂肪酸代謝に高度に濃縮された2つの代謝クラスターと有意に関連していた(Ganeshan and Chawla, 2014)。これらの結果は、マイクロバイオームが、腸内の免疫細胞との直接的な相互作用だけでなく、重要な代謝産物の全身的な利用可能性を調節するといった間接的なメカニズムによっても、免疫機能に多様な影響を及ぼす能力を有していることを浮き彫りにしている。
ここで我々は、抗生物質によるマイクロバイオームの擾乱が、健康な成人のワクチン接種に対する免疫応答に影響を及ぼす可能性を示した。抗生物質とワクチンは最も広く使用されている医療介入の2つであるため、このことは臨床診療と公衆衛生に重要な意味を持つ。これらの知見は、腸内細菌叢と我々の免疫系との相互作用を制御するメカニズムをよりよく理解しようとするさらなる研究に役立つはずである。
STAR★方法
主要資源表
試薬またはリソースソース識別子
抗体
抗ヒトCD19 FITC Beckman Coulter Cat #IM1284U ; RRID: AB_131011
抗ヒト CD27 PE Beckman Coulter Cat #IM2578 ; RRID: AB_131206
抗ヒト CD3 PerCP BD Biosciences Cat #340663 ; RRID: AB_400073
抗ヒト CD38 APC BD Biosciences Cat #340439 ; RRID: AB_400512
抗ヒト CD20 Pacific Blue Biolegend Cat #302328 ; RRID: AB_1595435
抗ヒト HLA-DR V500 BD Biosciences Cat #561224 ; RRID: AB_10563765
抗ヒト CD195(CCR5) BV605 BD バイオサイエンス Cat #563379 ; RRID: AB_2738167
抗ヒト CD138 PE-CF594 BD バイオサイエンス Cat #564606 ; RRID: AB_2738863
抗ヒト Ki-67 PE-CY7 BD バイオサイエンス Cat #561283 ; RRID: AB_10716060
抗ヒト CD3 Alexa Fluor 700 BD Biosciences Cat #557943 ; RRID: AB_396952
抗ヒト CD4 PerCP-Cy5.5 BD Biosciences Cat #341653 ; RRID: AB_400451
抗ヒト CD45RA BV605 Biolegend Cat #304134 ; RRID: AB_2563814
抗ヒト CD183(CXCR3) Alexa Fluor 488 BD Biosciences Cat #558047 ; RRID: AB_397008
抗ヒト CD196(CCR6) PE-CF594 BD バイオサイエンス Cat #564816 ; RRID: AB_2738971
抗ヒト CD279(PD-1) BV421 Biolegend Cat #329920 ; RRID: AB_10960742
抗ヒト CD14 APC-H7 BD Biosciences Cat #560180 ; RRID: AB_1645464
抗ヒト CD20 APC-H7 BD バイオサイエンス Cat #560734 ; RRID: AB_1727449
抗ヒト CD185(CXCR5) PE eBioscience Cat #12 -9185-42; RRID: AB_11219877
抗ヒト CD278(ICOS) Alexa Fluor 647 Biolegend Cat #313516 ; RRID: AB_2122582
抗ヒト Ki-67 PE-CY7 BD Biosciences Cat #561283 ; RRID: AB_10716060
抗ヒト CD3 FITC BioLegend Cat #344804 ; RRID: AB_2043993
抗ヒト CD19 FITC BioLegend Cat #302206 ; RRID: AB_314236
抗ヒト CD20 FITC BioLegend Cat #302304 ; RRID: AB_314252
抗ヒト CD56 FITC BioLegend Cat #318304 ; RRID: AB_604100
抗ヒト CD80 PE BioLegend Cat #305208 ; RRID: AB_314504
抗ヒト BDCA-1(CD1c) PerCP-Cy5.5 BioLegend Cat #331514 ; RRID: AB_1227535
抗ヒト CD123 PE-Cy7 BioLegend Cat #306010 ; RRID: AB_493576
抗ヒト CD33 BV421 BioLegend Cat #303415 ; RRID: AB_10900080
抗ヒト CD14 BV605 BioLegend Cat #301834 ; RRID: AB_2563798
抗ヒト CD11b BV650 BioLegend Cat #101239 ; RRID: AB_11125575
抗ヒト CCR7 BV711 BioLegend Cat #353227 ; RRID: AB_11219587
抗ヒト CD16 APC BioLegend Cat #302012 ; RRID: AB_314212
抗ヒト CD11c Alexa700 BD Cat #561352 ; RRID: AB_10612006
抗ヒト HLA-DR APC-Cy7 BD Cat #335814 ; RRID: AB_399991
抗ヒト CD21 FITC BioLegend Cat #354910 ; RRID: AB_2561575
抗ヒト CD38 PE BioLegend Cat #303505 ; RRID: AB_314357
抗ヒト CD19 PE-CF594 BD Cat #562294 ; RRID: AB_11154408
抗ヒト HLA-DR PerCP-Cy5.5 BioLegend Cat #307630 ; RRID: AB_893567
抗ヒト CD27 PE-Cy7 BioLegend Cat #356412 ; RRID: AB_2562258
抗ヒト IgD BV421 BD Cat #562518 ; RRID: AB_11153121
抗ヒト CD69 BV605 BioLegend Cat #310938 ; RRID: AB_2562307
抗ヒト CD20 BV650 BioLegend Cat #302336 ; RRID: AB_2563806
抗ヒト CD56 BV711 BD Cat #563169 ; RRID: AB_2738043
抗ヒト CD71 APC BioLegend Cat #334108 ; RRID: AB_10915138
抗ヒト CD16 Alexa700 BD Cat #560713 ; RRID: AB_1727430
抗ヒト CD3 APC-Cy7 BioLegend Cat #300318 ; RRID: AB_314054
抗ヒト IgA1-PE Southern Biotech Cat #9130 -09; RRID: AB_2796656
抗ヒト IgG1-PE Southern Biotech Cat #9052 -09; RRID: AB_2796621
抗ヒト IgG1 Southern Biotech Cat #9054 -01; RRID: AB_2796625
抗ヒトIgG2 Southern Biotech Cat #9070 -01; RRID: AB_2796636
抗マウス IgG Fc 特異的 Jackson Laboratories Cat #115 -035-071; RRID: AB_2338506
抗ヒト IgA HRP 標識 KPL Cat #14 -10-01
抗ヒト IgG HRP 標識 GE Healthcare Cat #NA933 ; RRID: AB_772208
抗ヒトIgG γ鎖特異的アルク-ホス結合体 Millipore Cat #401442 ; RRID: AB_437803
細菌およびウイルス株
H1N1 A/カリフォルニア/2009年7月 CDCより提供 N/A
H3N2 A/テキサス/50/2012 CDC提供 N/A
H3N2 A/スイス/9715293/2013 CDCより提供 N/A
B/マサチューセッツ/2/2012 CDC提供 N/A
B/プーケット/3073/2013 CDC提供 N/A
化学物質、ペプチド、組換えタンパク質
EDC ThermoFisher Cat #A35391
スルホ-NHS ThermoFisher Cat #24510
A/California/07/2009 HA タンパク質 Immune Technology Cat #IT -003-SW12p
A/California/07/2009 HAタンパク質 BEI Resources Cat #44074
A/California/07/2009 HAタンパク質 自家製 (Khurana et al., 2010) N/A
寄託データ
生データおよび処理済みデータ(遺伝子発現) 本論文 GEO: GSE120719
生データおよび処理済みデータ 本論文 ImmPort: SDY1086
生データ(16S rRNA遺伝子シーケンス) 本論文 SRA: PRJNA505336
ソフトウェアとアルゴリズム
Graphpad Prism v7 および 8 GraphPad Software https://www.graphpad.com
FlowJo v8, 9, 10 FlowJo https://www.flowjo.com/
Immunospot software v5 CTL http://www.immunospot.com/ImmunoSpot-analyzers-software
Bioconductor v3.5 https://www.bioconductor.org/
MATLAB R2015a MathWorks https://www.mathworks.com/products/matlab.html
GSEA v2.2.3 Subramanian et al., 2005 http://software.broadinstitute.org/gsea/index.jsp
apLCMS v6 Yu et al., 2009 http://web1.sph.emory.edu/apLCMS/
mummichog v2.0.6 Li et al., 2013 http://mummichog.org/
MMRN (hiconet v0.4.1) Li et al., 2017 https://github.com/shuzhao-li/hiconet
USEARCH v9.2 Edgar, 2010 https://www.drive5.com/usearch/
mothur v1.36 Schloss et al., 2009 https://www.mothur.org/
Cytoscape v3.5.1 Shannon et al., 2003 https://cytoscape.org
その他
カルボキシレート修飾マイクロスフェア Luminex Cat # varies
新しいタブで表を開く
リードの連絡先と材料の入手可能性
リソースおよび試薬に関する詳細情報およびリクエストは、リードコンタクトであるBali Pulendran (bpulend@stanford.edu)までご連絡ください。本研究では新規のユニークな試薬は作成していない。
実験モデルと被験者の詳細
2014-2015年および2015-2016年のシーズン中、合計33名の健康な成人を登録し、抗生物質投与群(n=16)と対照群(n=17)に無作為に割り付けた。被験者は、clinicaltrials.gov(NCT02154061)に記載されている適格基準を満たした18~40歳の男性および非妊娠女性であった。被験者のデモグラフィックを表S4に示す。抗生物質治療はネオマイシン、バンコマイシン、メトロニダゾールのカクテルを5日間経口投与した。抗生物質治療はワクチン接種日の3日前から開始され、抗生物質治療群ではその1日後まで続けられた。試験参加者は全員、2014-2015年シーズンまたは2015-2016年シーズンのFluzoneワクチン接種を受けた。各被験者から書面によるインフォームドコンセントを取得し、プロトコルはエモリー大学の施設審査委員会の承認を得た。
方法の詳細
治療プロトコールおよび検体収集
本試験はジョージア州アトランタで実施され、2期に分けて行われた。第1相では、2015年春に合計22人の被験者が登録され、抗生物質投与群(n=11)と対照群(n=11)に無作為に割り付けられた。2016年春に実施された第2相では、過去3回のインフルエンザシーズンにIIVを受けていない被験者を対象に、ワクチンに含まれる3種類のインフルエンザ株のうち少なくとも2種類について、MN力価<320の事前スクリーニングを行った。第2相試験では合計11人の被験者が登録され、抗生物質投与群(n=5)と対照群(n=6)に分けられた。1名(接種前の血液サンプルが得られなかった)を除くすべての被験者に3価インフルエンザワクチン(TIV)が接種された。ワクチンに含まれるインフルエンザ株を表S5に示す。抗生物質レジメンは、硫酸ネオマイシン(500mgを1日3回)、バンコマイシン(125mgを1日4回)、メトロニダゾール(500mgを1日3回)の3種類の抗生物質のカクテルで構成され、5日間経口投与された。ピルカウントは0日目、1日目、3日目に実施した。各被験者から試験実施前に書面によるインフォームド・コンセントを取得し、本研究はEmory University Institutional Review Boardにより承認された。血液と便のサンプルは、図1に示したように、一定の間隔で採取された。
細胞、血漿およびRNAの分離
末梢血単核球(PBMC)と血漿は、製造業者のプロトコールに従って新鮮血(CPTs; Vacutainer® with Sodium Citrate; BD)から分離した。PBMCは10%FBS添加DMSO中で凍結し、-80℃で保存した後、翌日液体窒素フリーザー(-196℃)に移した。CPTからの血漿サンプルは-80℃で保存した。新鮮なPBMCを溶解し(約1.5×106細胞に対して1mLのTrizol)、RNAを分解から保護するためにTrizol(Invitrogen)を用いた。Trizolサンプルは-80℃で保存した。
マイクロバイオーム解析
サンプルの分離
Second Genome社(https://www.secondgenome.com)は、MoBio PowerMag® Microbiome kit(カリフォルニア州カールスバッド)を用いて、メーカーのガイドラインに従い、ハイスループット処理に最適化された糞便サンプルから核酸単離を行った。すべてのサンプルは、Qubit® Quant-iT dsDNA High Sensitivity Kit(Invitrogen, Life Technologies, Grand Island, NY)を用いて定量し、DNAの最小濃度と質量を満たしていることを確認した。
ライブラリー調製
バクテリア16S V4 rDNA領域のサンプルを濃縮するために、イルミナ(カリフォルニア州サンディエゴ)のアダプターおよびインデキシングバーコードを組み込んだ配列でテーリングされた周辺の保存領域に対してデザインされた融合プライマーを用いてDNAを増幅した。各サンプルは、2つの異なるバーコード付きV4融合プライマーでPCR増幅された。PCR後の定量下限を満たし、プールとシーケンスに進められたサンプル。各サンプルについて、PCR産物を精製するための固相可逆固定化法を用いて増幅産物を濃縮し、qPCRで定量した。
プロファイリング法
16S V4に濃縮され、増幅され、バーコード化されたサンプルを含むプールをMiSeq®試薬カートリッジにロードし、フローセルとともに装置にセットした。MiSeq装置上でクラスター形成後、ペアエンドシーケンス用に設計されたカスタムプライマーでアンプリコンを250サイクルシーケンスした。
qPCRアッセイ
菌株の定量は、ABI 5900htのSYBRGreenを用いたqPCRで行った。ユニバーサル16Sプライマーを使用した:
16su-f 5′-actcctacgggaggcagcagt-3′。
16su-R1 5′-tattaccgcggctgctggc-3′ - 16su-R1 5′-tattaccgcggctgctggc-3′
反応は10μLのSYBRGreen MasterMix (2x, VeriQuest)、1μLの各プライマー(10uM)、6μLのPCRグレードの水、および2μLのDNA鋳型(<100ng)から構成された。反応条件は、50℃で2分、95℃で10分、95℃で15秒、54℃で30秒、72℃で30秒のサイクルを40回繰り返した。各サンプルは3連で行い、平均値を用いた。各qPCRアッセイで標準曲線を作成し、サンプルの定量を決定するために使用した。サンプルあたりの16Sコピーの平均量は、便1gあたりの標的遺伝子コピーで正規化した。
フラジェリンとLPSの定量
糞便中の生理活性フラジェリンおよびリポ多糖(LPS)の定量は、それぞれヒト胚性腎臓(HEK)-blue-mTLR5細胞およびHEK-blue-mTLR4細胞(Invivogen, San Diego, CA, USA)を用いて既述のように行った(Chassaing et al.) 糞便サンプルを100 mg/mLの濃度に調製し、ホモジナイズしてから8000 gで10分間遠心分離し、得られた上清を連続希釈して細胞株で測定した。HEK-Blue-mTLR5細胞およびHEK-Blue-mTLR4細胞を用いた標準曲線測定には、それぞれ精製ネイティブフラジェリン(Salmonella Typhimurium)およびLPS(Sigma-Aldrich)を用いた。血漿中のフラジェリンを測定するために、20μlの希釈血漿(1:5)を180μlの培養液に添加した。24時間の刺激後、細胞培養上清をQUANTI-Blue培地(Invivogen)にアプライし、アルカリホスファターゼ活性を640nmで測定した。
LPS特異的IgAおよびIgG抗体の測定
LPS特異的IgAおよびIgGレベルはELISAで測定した。マイクロタイタープレートに精製LPS(2μg/ウェル)をコートした。被験者の血漿サンプルを1:200に希釈した後、コーティングしたプレートに塗布した。インキュベーションおよび洗浄後、ウェルを西洋ワサビペルオキシダーゼと結合した抗ヒトIgA(KPL)またはIgG(GEヘルスケア)とインキュベートした。比色ペルオキシダーゼ基質テトラメチルベンジジンを用いて血漿中の総IgAおよびIgGを定量し、ELISAプレートリーダーを用いて各ウェルの光学濃度(OD)を450nmで読み取った。
インフルエンザマイクロ中和アッセイ
ウイルス中和活性は、疾病対策予防センター(CDC)のパンデミックインフルエンザ基準研究所の方法に基づき、MDCK細胞を用いたマイクロ中和(MN)アッセイで分析した。H1N1-A/カリフォルニア/7/2009、H3N2-A/テキサス/50/2012、H3N2-A/スイス/9715293/2013、B/マサチューセッツ/2/2012およびB/プーケット/3073/2013の季節性インフルエンザワクチン株に対するMNによる抗体中和価を測定した。血清は1:20の初期希釈で検査され、陰性(1:20未満)のものは力価10とされた。すべての血清は3連で検査し、幾何平均値を解析に用いた。
ELISAによる抗原特異的IgG1およびIgG2抗体の測定
ヘマグルチニン特異的ELISAを用いて、ヒト血清中のIgGサブクラスを測定した。Thermo 2 HB Immulon Microtiter Plates Flat-Bottom (supplier no. 3355)にA/California/07/09 HA0 protein (BEI Resources)を一晩コートした。プレートを洗浄し、Novex Human IgG Subclass Profile Novex (Life Technologies)キットのThermo希釈液を用いてブロックした。血清希釈液(1:500、1:1000、1:2000、1:4000)をコートしたプレートに加え、1時間静置した。洗浄後、抗ヒトIgG1またはIgG2 [Southern Biotech mouse anti-human IgG1 (9054-01)およびIgG2 (9070-01)]を添加し、1時間静置した。洗浄後、Goat anti-mouse IgG Fc-specific Affinipure(Jackson 115-035-071)を添加し、Thermo TMB溶液で反応性を確認した。プレートは Molecular Devices 社のプレートリーダーで読み取った。
Luminex による抗原特異的 IgG1 抗体および IgA1 抗体の測定
抗原特異的抗体のサブクラス/アイソタイプは、ハイスループットのLuminexベースのアッセイを用いて決定した(Brown et al.) H1 A/California/07/2009抗原はImmune Technology Corp. この抗原は、EDC(ThermoFisher)および Sulfo-NHS(ThermoFisher)を介した共有結合 NHS-エステル結合により、カルボキシレート修飾マイクロスフェア(Luminex Corp. この抗原コートマイクロスフェアを非結合性 384 ウェルプレート(Grenier Bio-One、Kremsmunster、 Austria)に 1 ウェルあたり 1000 ビーズ(45 μl)添加した。血清サンプルはIgG1についてはPBSで1:100に希釈し、他のサブクラスについてはビーズとインキュベートする前に1:10に希釈した。希釈した血清検体5μLをミクロスフェアの入ったプレートに加え、4℃で一晩振とう器でインキュベートした。ミクロスフェアを0.1% BSAと0.05% Tween-20を含むPBS緩衝液で洗浄し、二次検出器、すなわちPE標識抗IgG1または-IgA1検出抗体(Southern Biotech, Birmingham, AL)と共に室温で1時間インキュベートした。マイクロスフェアーを前と同様に洗浄し、iQue Screener Plus (Intellicyt Corp., Albuquerque, NM)で読み取った。
表面プラズモン共鳴によるポリクローナル血清抗体のrHAタンパク質への結合動態と抗体親和性
ワクチン接種後の個々の血清の定常状態平衡結合は、前述したようにProteOn表面プラズモン共鳴バイオセンサー(BioRad)を用いて25℃でモニターした(Khurana et al.) 自社で調製したH1N1-A/California/7/2009由来のrHAタンパク質(Khurana et al., 2010)を、テストフローセル内で500共鳴単位(RU)を持つアミンカップリング付きGLCセンサーチップに結合させた。10倍および100倍に希釈した60μLの新鮮な血清サンプルを50μL/分の流速で注入し(接触時間120秒)、会合させ、1200秒のインターバルで解離させた(流速50μL/分)。タンパク質表面からの反応は、モック表面からの反応と、別のバッファーのみの注入からの反応について補正した。 mAb 2D7(抗CCR5)は、これらの実験でネガティブコントロールとして使用した。全抗体結合は、前述したように、BioRad ProteOn managerソフトウェアを用いたSPRにより、血清サンプルとインフルエンザウイルスのrHAタンパク質との相互作用から直接決定した。抗体オフレート定数は、1秒間に減衰する抗原抗体複合体の割合を表す定数で、解離期のSPRを用いて血清/血漿サンプルとrHAの相互作用から直接決定し、前述のように不均一サンプルモデルに対してBioRad ProteOn managerソフトウェアを用いて計算した。オフレート定数は2回の独立したSPR測定から求めた。
免疫集団のフローサイトメトリー解析
フローサイトメトリー解析は、ヘパリン処理した全血(形質芽細胞および血球濾胞Tヘルパー様細胞)または凍結保存したPBMC(活性化B細胞および自然免疫集団)のいずれかで行った。
全血
全血アッセイでは、まず細胞を適切な抗体カクテルで室温で20分間表面染色した後、BD FACS lysing solution(BD Biosciences)を用いて赤血球溶解を行った。その後、細胞をBD Cytofix/Cytopermで氷上30分間固定・透過処理し、抗Ki67 PE-CY7(BD Biosciences)をBD透過処理バッファー中で氷上30分間添加した。全サンプルはLSR II(BD Biosciences社製)で測定し、FlowJoソフトウェア(FlowJo, LLC社製)を用いて解析した。
形質芽細胞の同定のために、細胞を抗CD19 FITC、抗CD27 PE、抗CD3 PerCP、抗CD38 APC、抗CD20 Pacific Blue、抗HLA-DR V500、抗CD195 BV605、抗CD138 PE-CF594抗体のカクテルで染色した。形質芽細胞はCD3-、CD19+、CD27++、CD38++細胞として同定され、全CD19+ B細胞に対する割合として定量された。
血中Tfh様細胞の同定には、抗CD3 Alexa Fluor 700、抗CD4 PerCP-Cy5.5、抗CD45RA BV605、抗CD183 Alexa Fluor 488、抗CD196 PE-CF594、抗CD279 BV421、抗CD14 APC-H7、抗CD20 APC-H7、抗CD185 PE、および抗CD278 Alexa Fluor 647抗体のカクテルを用いた。活性化血中Tfh1型細胞は、Schmitt and Ueno, 2013により以前に記載されている。循環Tfh様細胞のこのサブセットは、我々の研究ではCD3+、CD4+、CXCR5+、CXCR3+、CCR6-、ICOS+、PD-1+と定義され、ワクチン接種後7日目の頻度レベルは、0日目の頻度に対するlog2倍変化として示された。
凍結保存PBMC
2-3×106個のPBMCを解凍し、適切な抗体カクテルを用いて室温で20分間染色した。細胞は5%ウシ胎児血清を含むPBSで洗浄し、Cytofix buffer(BD)で固定した後、LSR IIフローサイトメーター(BD Biosciences)で解析した。すべてのフローサイトメトリー解析は、FlowJoソフトウェア(FlowJo, LLC)を用いて行った。
活性化/非活性化骨髄系(mDC)および形質細胞系(pDC)樹状細胞の同定には、抗CD3 FITC、抗CD19 FITC、抗CD20 FITC、抗CD56 FITC、抗CD80 PE、抗BDCA-1 PerCP-Cy5.5のカクテルを用いた。 5、抗CD123 PE-Cy7、抗CD33 BV421、抗CD14-BV605、抗CD11b BV650、抗CCR7 BV711、抗CD16 APC、抗CD11c Alexa700、HLA-DR APC-Cy7抗体が使用された。活性化された(CD80+)CD16+ mDCは、CD3-CD19-CD20-CD56-細胞としてシングルツゲート内で定義され、CD14- HLA-DR+ CD123- CD11c+ CD16+ CD1c- CD80+として一意に同定された。活性化pDCもCD80+である。
活性化B細胞(ABC)の同定には、抗CD21 FITC、抗CD38 PE、抗CD19 PE-CF594、抗HLA-DR PerCP-Cy5.5、抗CD27 PE-Cy7、抗IgD BV421、抗CD69 BV605、抗CD20 BV650、抗CD56 BV711、抗CD71 APC、抗CD16 Alexa700、および抗CD3 APC-Cy7抗体のカクテルを用いた。ABCはEllebedyら、2016により以前に記載された。ABCはCD3-CD19+CD20+IgD-CD71+として一意に同定された。
ワクチン特異的抗体分泌細胞(ASC)のELISPOT測定
ELISPOTプレート(Millipore, cat.#MAIPS4510)に、PBS中9μg/mlの3価インフルエンザワクチン(Fluzone)を4℃で一晩コートし、その後洗浄し、非特異的結合を避けるために完全培地を37℃で2時間添加した。翌日、IgGを分泌するワクチン特異的形質芽細胞の数を測定するため、洗浄したPBMCの希釈液(6x105から2x104細胞の範囲)をELISPOTプレートに加え、37℃、5%CO2で一晩インキュベートした。インキュベーション後、プレートを洗浄し、アルカリホスファターゼ標識ヤギ抗ヒトIgG抗体(ミリポア)を室温(RT)で4時間添加した。その後、アルカリホスファターゼ(AP)基質(Bio-Rad、cat. #170 -6432)を各ウェルに添加し、スポットを明らかにした。プレートをスキャンし、CTL Immunospotリーダーソフトウェアを用いてスポットをカウントした。
インフルエンザ反応性hmAbsの体細胞超変異(SHM)解析
ワクチン接種7日後に形質芽細胞(CD3-CD19+CD27hiCD38hi)を96ウェルPCRプレート(Bio-Rad)に単一細胞選別した。モノクローナル抗体配列を、可変重鎖および軽鎖遺伝子の単細胞PCR増幅によって増幅し、ヒトIgG発現ベクターに、以前に記載されたように挿入した(Chenら、2018、Henry Dunandら、2016、Wrammertら、2008)。 mAbのそれぞれの個々の重鎖および軽鎖発現ベクターをHEK293T細胞にトランスフェクトすることによってmAbを作製し、トランスフェクションの6日後にプロテインAアガロースビーズを用いてmAbを精製した。精製されたmAbは、季節性ワクチンとの結合、および季節性ワクチンの個々のウイルス成分との結合について試験された。SHMデータは、IgBlastに登録されている生殖細胞系列の配列とmAbの配列を比較することにより得られた。
マイクロアレイ実験
新鮮なPBMC(約1.5×106細胞)からの全RNAは、製造者の指示に従ってTrizol®(Invitrogen, Life Technologies Corporation)を用いて精製した。すべてのRNAサンプルは、ND-1000 spectrophotometer(NanoDrop Technologies社製)を用いて純度をチェックし、2100 BioAnalyzer(Agilent Technologies社製)を用いて電気泳動により完全性をチェックした。Affymetrix 社のプロトコールに従い、1 サンプルあたり 50 ng のインタクト RNA を用いて、2 ラウンドの in vitro 転写増幅と標識が行われた。Human U133 Plus 2.0 Arrays(GeneTitanプラットフォーム、Affymetrix社製、または個々のカートリッジを使用)をHybridization Oven 640(Affymetrix社製)を用いて45℃、60rpmで16時間ハイブリダイゼーションした後、スライドを洗浄し、Fluidics Station 450(Affymetrix社製)で染色した。スキャンは第7世代GeneChip Scanner 3000(Affymetrix)で行い、Affymetrix GCOSソフトウェアを用いて画像解析を行い、生の強度データを作成した。
血漿メタボローム解析
メタボロミクス解析は、以前に記載されたものと同様に行った(Li et al.) 簡単に説明すると、血漿65 mLにアセトニトリル130 mLを加え、混合した後、氷上で30分間インキュベートし、10分間遠心分離(4℃で13,400 x rpm)して代謝物を抽出した。上清をLC-MS分析用のオートサンプラーバイアルに移した。質量分析データは、Thermo Q Exactive質量分析計(Thermo Fisher, San Diego, CA)で、質量電荷比(m/z)85から2000までを記録するように設定し、10分間の逆相グラジエントで収集した。正負両方のエレクトロスプレーイオン化を用いた逆相クロマトグラフィー(Higgins Analytical社製、Targa C18 5 mM、100 3 2.1 mm)を用いて、各サンプルについて3テクニカルレプリケートを実施した。
液体クロマトグラフィー質量分析(LC-MS)による胆汁酸の測定
胆汁酸の測定は、以前に記載された方法(Voelker et al. 簡単に説明すると、ヒト血漿中の一次および二次胆汁酸濃度は、Thermo Q Exactive Focus質量分析計と組み合わせた島津CBM-20A HPLCシステムを用いて定量した。クロマトグラフィーには、Phenomenex C18カラム(4.6×100mm 2.6μm)を使用した。移動相Aは水中10mM酢酸アンモニウム、移動相Bは50:50メタノール:アセトニトリル(v/v)であった。カラム温度は60℃に保たれ、グラジエントは35%の移動相Bで2分間保たれた後、4.5分で55%の移動相Bに上げられた。その後、グラジエントは55%移動相Bで2分間保持した。最後に95%移動相Bで1分間溶出した。流速は1.2mL/分であった。質量分析には、Thermo Scientific Q Exactive Focus質量分析計(HESI搭載、負イオンモード)を使用した。イオン源の温度は400℃に設定した。定量は、分解能70,000のターゲットSIMを用いて行った。使用した標準物質は、CDCA(チェノデオキシコール酸)、DCA(デオキシコール酸)、gCDCA(グリコチェノデオキシコール酸)、gDCA(グリコデオキシコール酸)、tCDCA(タウロチェノデオキシコール酸)、 tDCA(タウロデオキシコール酸)、gLCA(グリコールリトコール酸)、CA(コール酸)、tCA(タウロコール酸)、tLCA(タウロリトコール酸)、LCA(リトコール酸)、gCA(グリココール酸)。gCDCAについては、標準品をアセトニトリルに溶解し、2.5ng/mL、5ng/mL、10ng/mL、50ng/mL、200ng/mL、500ng/mL、1500ng/mL、2250ng/mL、2500ng/mL、5000ng/mLの10種類の濃度勾配を作成した。他の11種類の胆汁酸については、標準物質をアセトニトリルに溶解し、0.5ng/mL、1ng/mL、2ng/mL、10ng/mL、40ng/mL、100ng/mL、300ng/mL、450ng/mL、500ng/mL、1000ng/mLの10種類の濃度勾配を作成した。
定量化と統計解析
マイクロバイオームデータ解析
OTU選択
配列決定されたペアエンドリードはUSEARCH (Edgar, 2010)を用いてマージされ、得られた配列はUSEARCH (usearch_global)を用いて社内の菌株データベースと比較された。同一性が99%以上のユニークな菌株にヒットしたすべての配列に、菌株のOperation Taxonomic Unit (OTU)を割り当てた。株ヒットの特異性を確保するため、最もよくヒットした株の同一性と2番目によくヒットした株の同一性の間に > = 0.25%の差が必要であった(例えば、99.75対99.5)。各菌株OTUについて、マッチするリードの1つを代表として選択し、すべての配列を菌株OTU代表に対してUSCEARCH(usearch_global)でマッピングし、菌株存在量を計算した。残りの非菌株配列は、USEARCHで品質フィルターされ、重複除去された。得られたユニーク配列は、UPARSE(de novo OTU clustering)により97%でクラスタリングされ、de novo OTUごとに代表的なコンセンサス配列が決定された。UPARSEクラスタリングアルゴリズムはキメラフィルタリングで構成され、キメラの可能性が高いOTUは除外される。品質フィルタリングに合格したすべての非菌株配列は、de novo OTUの存在量表を作成するために代表的なコンセンサス配列にマップされた。代表OTU配列は、99%でクラスタ化された16S rRNA遺伝子配列のGreengenes参照データベースに対してトレーニングされたmothur (Schloss et al., 2009)のベイジアン分類器によって分類学的分類が割り当てられた。
アルファ多様性(サンプル内多様性)メトリクス
観察された」多様性は、各サンプルで見つかったユニークなOTUの単純な合計であり、サンプルの豊かさとしても知られている。シャノン多様性(H)は、サンプルの豊かさと存在するOTUの相対的な存在量を利用して、以下の式に従って多様性指数を計算する:
=
∑
�
1
�
�
�
ln
�
�
ここで、piはi番目のOTUの相対存在量、SはOTUの総数である。
ベータ多様性(サンプル間の非類似度)メトリクス
すべてのプロファイルは、非類似度スコアを決定し、距離非類似度行列に格納するために、ペアワイズ方式で相互比較される。類似サンプルを比較する場合、距離関数は低い非類似度スコアを生成します。Bray-Curtis非類似度を用いて、豊度重み付けされたサンプルのペアワイズ差を計算した。Bray-Curtis非類似度は、2つのサンプルの存在量の合計に対するカウントの絶対差の合計の比によって計算される(Bray and Curtis, 1957)。2値の非類似度はJaccard indexで計算した。この指標は、少なくとも一方のサンプルに存在するOTUの数に対する、2つのサンプルのミスマッチ(一方には存在するが、他方には存在しないOTU)の数を比較するものである(Jaccard, 1912)。
順序付け
主座標分析(PCoA)(Gower, 1966)を用いて2次元の順序付けを行い、サンプル間の関係をグラフ化した。PCoAは、標本間の複雑な関係を視覚化するのに使用される2次元順序プロットの方法である。PCoAでは、サンプル間の非類似度値を用いて、非類似度値とプロット距離の線形相関を最大化することで、点を相対的に配置する。
トランスクリプトミクスデータ処理とパスウェイ/モジュール解析
初期データの品質は、バックグラウンドレベル、3′標識バイアス、およびサンプル間のペアワイズ相関によって評価した。異常値サンプルの CEL ファイルは除外し、同一試験に属する全サンプルの残りの CEL ファイルをグループ化し、Bioconductor で RMA (Irizarry et al., 2003) により正規化した。複数の遺伝子にマッピングされたプローブは破棄され、残りのプローブは、両試験の全被験者にわたって平均発現量が最も高い各遺伝子のプローブを選択することにより、遺伝子レベルに折りたたまれた。統計検定と相関分析はMATLABを用いて行った。検定の詳細と有意性のカッ トオフは図中の凡例に記載されている。BTMの濃縮検定はGSEA (Subramanian et al., 2005)を用いて行った。BTMはCytoscape (Shannon et al., 2003)を用いて可視化した。
メタボローム解析とパスウェイ解析
メタボロミクスソフトウェアパッケージapLCMS (Yu et al., 2009)を用いて代謝物ピークを抽出した。50%以上のサンプルに存在するピークのみを解析に使用した。分析品質の指標として、3つのテクニカルレプリケート間のピアソン相関係数Rを使用した。3つのテクニカルレプリケート間の一対相関がR < 0.7である場合、サンプルは除去された。そうでない場合は、テクニカルレプリケートの平均値を使用した。1つのテクニカルレプリケートが他の2つのテクニカルレプリケートとR < 0.7であった場合、それは取り除かれ、他の2つが平均された。各サンプルについて、データはlog2変換された。正イオンモードで5572ピーク、負イオンモードで5745ピークが検出された。統計検定、相関分析、PCAはMATLABを用いて行った。検定の詳細と有意性のカットオフは、図中の凡例に記載されている。Mummichogソフトウェア(Li et al., 2013)を使用して、各時点とベースライン(-21日目または0日目)の間の一対のt検定によってp < 0.05として選択された有意なピークを使用して、代謝経路における濃縮を同定した。
MMRNの構築とクエリー
トランスクリプトミクス、メタボロミクス、マイクロバイオミクス、および細胞頻度のMMRN統合は、更新されたソフトウェア(hiconet)を使用して、以前に記載された(Li et al.) 簡単に説明すると、まず、コミュニティ検出のための一般的なモジュール性アルゴリズムであるLeidenアルゴリズム(Traag et al. トランスクリプトミクスデータについては、クラスタリングの前に追加のステップを実施し、BTMに折りたたみ、モジュール活性スコアをメンバー遺伝子の平均値とした。メタボロミクスデータは、前述のように、クロマトグラフィーの保持時間を考慮してLC-MSデータ用にカスタマイズした階層的クラスタリング法を用いてクラスタリングした(Gardinassi et al.) 次に、各データタイプの特徴クラスター間の関連を、部分最小二乗(PLS)回帰によって推定した。関連性の有意性は、特徴とサンプルラベルの両方を再サンプリングした並べ替えデータと比較することで計算した。得られたネットワークは、Cytoscape (Shannon et al., 2003)を用いて可視化した。LCAまたはH1N1特異的IgG1データとの関連についてネットワークを照会するために、すべてのネットワーク特徴(BTM、m/zピーク、OTU、および細胞頻度)のランク付けされたリストが、ランク付け指標としてLCAまたはIgG1のいずれかとのスピアマン相関を用いて作成された。その後、MMRN内の各クラスタを「遺伝子セット」として扱い、GSEA (Subramanian et al., 2005)を用いて濃縮検定を行い、ランク付けされたリストのどちらかの端でメンバーフィーチャーが濃縮されている(LCAまたはIgG1と高い相関がある)クラスタを同定した。
データとコードの利用可能性
この研究で作成されたデータは、ImmPort (https://www.immport.org/shared/home)のアクセッション番号ImmPort: SDY1086で入手可能である。また、マイクロアレイデータはGEO: マイクロアレイデータはGSE120719で、16S rRNAシーケンスデータはSRA: PRJNA505336で入手できる。
その他のリソース
臨床試験登録 (NCT02154061): https://clinicaltrials.gov/ct2/show/NCT02154061
謝辞
臨床試験に協力してくれたHope Clinicのスタッフ、特にMary Bower、Eileen Osinski、Briyana Domjahnに感謝する。また、マイクロバイオームデータの作成と解析に携わったLuisa Chan、Kathryn Iverson、Second Genome、LC-MSによる胆汁酸の測定に携わったLily ChenとCreative Proteomics、データ管理とキュレーションに携わったDmitri Kazmin、技術支援に携わったChris IbegbuとEmory Immunology/Flow Cytometry Core、ライブラリーの調製に携わったAmber N. Wohlabaugh:ライブラリー調製と配列決定、Mark MulliganとAnesh Mehta:臨床試験デザインに関する助言、Mark Davis、Rafi Ahmed、David Relman、Justin Sonneburg、Taia Wang:有益な議論。本研究は、Emory-UGA CEIRS契約HHSN272201400004C(B.P.および研究代表者Walt Orensteinへ)、NIH助成金HIPC U19AI090023(B.P.へ)、U19AI057266(B.P.および研究代表者Rafi Ahmedへ)の支援を受けた。および研究責任者Rafi Ahmed)、UH2AI132345(S.L.へ)、U24AI120134(S.E.B.)、T32AI07290(T.H.へ)、Sean Parker Cancer Institute、Soffer基金(B.P.)、Violetta Horton基金(B.P.)。
著者貢献
独創的プロジェクトの構想および立案、B.P.およびN.R.、プロジェクト全体を通しての知的貢献、B.P.、N.R.、T.H.およびM.C.、形式的分析、T.H.、M.C.、J.D.、 A.A.U.、L.G.、調査、M.C.、C.B.、C.L.、M.S.M.、H.W.、S.K.、J.G.、N.-Y.Z.、M.Huang、C.P.、K.G.、B.C.、J.Z.、A.B.、M.Hahn、可視化、M.C.、T.H.、検証、C.B、 C.L.、G.A.;執筆-原案、T.H.、M.C.、B.P.;執筆-校閲・編集、T.H.、M.C.、B.P.;監修、B.P.、N.R.、A.T.G.、S.E.B.、P.C.W.、S.K、 H.G.、S.L.、G.A.、プロジェクト管理、M.C.、リソース、M.P.M.、S.J.J.、S.K.、H.G.、S.L.、G.A.、A.T.G.、P.C.W.、S.E.B.、資金獲得、B.P.、S.L.、S.E.B.、T.H.。
利害関係の宣言
著者らは、競合する利益はないと宣言している。
補足情報
pdfをダウンロード(.1 MB)
pdfファイルのヘルプ
資料S1. 表S1、S2、S4、S5
xlsxをダウンロード (.11 MB)
xlsxファイルのヘルプ
表S3. 図7に関連するMMRNのノードのメンバーの特徴とパスウェイの濃縮度
参考文献
Agus A.
Planchais J.
Sokol H.
健康および疾患におけるトリプトファン代謝の腸内細菌叢制御。
Cell Host Microbe. 2018; 23: 716-724
論文で見る
スコープス (1122)
PubMed
要旨
全文
全文PDF
グーグル奨学生
バッケD.
Sun J.
新しい機能を持つ古代の核内受容体VDR: マイクロバイオームと炎症。
炎症。Bowel Dis. 2018; 24: 1149-1154
論文で見る
スコープス (83)
PubMed
クロス
グーグル奨学生
ベカッティーニ S.
タウル Y.
パマー E.G.
抗生物質による腸内細菌叢の変化と疾患。
トレンドMol. Med. 2016; 22: 458-478
論文で見る
スコープス (532)
PubMed
概要
全文
全文PDF
グーグル奨学生
ベルカイド Y.
ハンドT.W.
免疫と炎症における微生物叢の役割。
Cell. 2014; 157: 121-141
論文で見る
スコープス (2904)
PubMed
概要
全文
全文PDF
グーグル・スカラー
ブレイ J.R.
カーティス J.T.
ウィスコンシン州南部の高地林群集の序列化。
Ecol. 1957年; 27: 326-349
論文で見る
クロス
グーグル・スカラー
ブレントリー J.M.
プライスD.A.
シャッカー T.W.
アッシャー・T.E.
シルベストリG.
ラオ S.
カザズ Z.
ボーンスタイン E.
ランボットO.
アルトマンD.
他。
慢性HIV感染症における全身性免疫活性化の原因である微生物転位。
Nat. Med. 2006; 12: 1365-1371
論文で見る
スコパス(2785)
PubMed
クロス
グーグル奨学生
ブロディン P.
ヨイッチV.
ガオ T.
バッタチャリヤ S.
エンジェルC.J.
ファーマン D.
シェン・オアー S.
デッカー C.L.
スワン G.E.
ビュートA.J.
他
ヒトの免疫系における変異は、遺伝的でない影響によって大きく左右される。
Cell. 2015; 160: 37-47
論文で見る
スコープス (658)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ブラウン E.P.
リヒト A.F.
デュガスト A.S.
Choi I.
ベイリー-ケロッグC.
アルター G.
アッカーマン M.E.
臨床検体からの抗原特異的抗体の高スループット多重IgGサブクラス分類。
J. Immunol. Methods. 2012; 386: 117-123
論文で見る
スコープス (144)
PubMed
クロス
グーグル奨学生
ケーヒル C.M.
ロジャースJ.T.
インターロイキン(IL)1βによるIL-6の誘導は、アクチベーター・プロテイン-1を標的とする新規のホスファチジルイノシトール3キナーゼ依存性AKT/IkappaBキナーゼα経路によって媒介される。
J. Biol. Chem. 2008; 283: 25900-25912
論文で見る
スコープス (180)
PubMed
クロス
グーグル奨学生
カラボッティ M.
シロッコA.
マセッリ M.A.
セヴェリC.
腸脳軸:腸内細菌叢、中枢および腸神経系間の相互作用。
アン。Gastroenterol. 2015; 28: 203-209
論文で見る
PubMed
グーグル奨学生
シャセインB.
コレンO.
グッドリッチJ.K.
プール A.C.
スリニヴァサン S.
レイ・R.E.
ゲヴィルツ A.T.
食事性乳化剤は、大腸炎とメタボリックシンドロームを促進するマウスの腸内細菌叢に影響を与える。
Nature. 2015; 519: 92-96
論文で見る
スコープス (1246)
PubMed
クロス
グーグル奨学生
チェン Y.Q.
ヴォールボルドT.J.
Zheng N.Y.
Huang M.
Huang Y.
Neu K.E.
リー J.
ワン H.
ロハス K.T.
カークパトリックE.
ほか
ヒトにおけるインフルエンザ感染は、広範な交差反応性および保護的ノイラミニダーゼ反応性抗体を誘導する。
Cell. 2018; 173: 417-429
論文で見る
スコープス (241)
PubMed
要旨
全文
全文PDF
グーグル奨学生
コリンズ N.
ベルカイド Y.
微生物叢はワクチンや病原体に対する防御免疫に影響するか?内因性アジュバントの関与。
Cold Spring Harb. Perspect. Biol. 2018; 10 (a028860)
論文で見る
スコープス (20)
PubMed
クロス
グーグル奨学生
Crotty S.
T濾胞ヘルパー細胞の分化、機能、および疾患における役割。
Immunity. 2014; 41: 529-542
論文で見る
スコパス (1237)
PubMed
要旨
全文
全文PDF
グーグル奨学生
デスレフセンL.
レルマンD.A.
抗生物質の反復擾乱に対するヒト遠位腸内細菌叢の不完全な回復と個体差反応。
Proc. Natl. Sci. USA. 2011; 108: 4554-4561
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
ドミンゲス-ベロ M.G.
コステロE.K.
コントレラス M.
マグリス M.
イダルゴ G.
フィーラー N.
ナイトR.
分娩様式は、新生児の複数の身体生息地にわたる初期微生物叢の獲得と構造を形成する。
Proc. Natl. Sci. USA. 2010; 107: 11971-11975
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
Edgar R.C.
BLASTよりも桁違いに高速な検索とクラスタリング。
Bioinformatics. 2010; 26: 2460-2461
論文で見る
筑波大学
PubMed
クロスリファレンス
グーグル奨学生
エレベディ A.H.
ジャクソンK.J.
キシック H.T.
ナカヤ H.I.
デイビス C.W.
ロスキン K.M.
マッケロイ A.K.
オシャンスキー C.M.
エルバイン R.
トーマス S.
他
ウイルス感染またはワクチン接種後のヒト血液における抗原特異的形質芽細胞および記憶B細胞サブセットの定義。
Nat. Immunol. 2016; 17: 1226-1234
論文で見る
スコープス (240)
PubMed
クロス
グーグル奨学生
フラスカD.
ディアス A.
ロメロ M.
メンデス N.V.
ランディン A.M.
Blomberg B.B.
2011-2012年インフルエンザワクチンシーズンに評価されたH1N1特異的血清IgG1およびIgG3レベルに対する年齢の影響。
免疫。Ageing. 2013; 10: 14
論文で見る
スコープス (53)
PubMed
クロス
グーグル奨学生
フロムヘルツL.
バブA.
フンメルE.
リスト M.J.
ロート A.
ワツル B.
クーリング S.E.
健康成人における血漿胆汁酸濃度の加齢変化--横断KarMeN研究の結果。
PLoS ONE. 2016; 11: e0153959
論文で見る
スコープス (57)
PubMed
クロス
グーグル奨学生
ガネーシャン K.
チャウラ A.
免疫応答の代謝制御。
Annu. Rev. Immunol. 2014; 32: 609-634
論文で見る
スコパス(531)
PubMed
クロス
グーグル奨学生
ガルディナッシ L.G.
アレバロ-エレラM.
エレーラ S.
コーディ R.J.
トラン V.
スミス M.R.
ジョンソン M.S.
チャッコ B.
リュー K.H.
ダーリー-ウスマーV.M.
他
MaHPICコンソーシアム
原虫に対する臨床耐性の統合的メタボロミクスおよびトランスクリプトミクスシグネチャーは、自然細胞免疫およびT細胞シグナル伝達の活性化を明らかにした。
レドックスバイオロジー 2018; 17: 158-170
論文で見る
スコープス (47)
PubMed
クロス
グーグル奨学生
ジオンゴ A.
ガノ K.A.
クラッブ D.B.
ムカルジー N.
ノベロ L.L.
カセラ G.
ドリュー J.C.
イロネン J.
クニップ M.
Hyöty H.
他。
1型糖尿病の自己免疫マイクロバイオームの定義に向けて。
ISME J. 2011; 5: 82-91
論文で見る
スコープス (623)
PubMed
クロス
グーグル奨学生
ゴパラクリシュナンV.
スペンサーC.N.
ネジ・L.
ルーベン A.
アンドリュース M.C.
カルピネッツT.V.
プリエト P.A.
ビセンテ D.
ホフマン K.
ウェイ S.C.
et al.
腸内細菌叢はメラノーマ患者の抗PD-1免疫療法に対する反応を調節する。
Science. 2018; 359: 97-103
論文で見る
スコープス (2607)
PubMed
クロス
グーグル奨学生
Gower J.C.
多変量解析に用いられる潜在根とベクトル法の距離特性.
Biometrika. 1966; 53: 325
論文で見る
クロス
グーグル奨学生
郭 C.
Xie S.
Chi Z.
Zhang J.
Liu Y.
Zhang L.
Zheng M.
Zhang X.
Xia D.
Ke Y.
et al.
胆汁酸はNLRP3インフラマソームの阻害を介して炎症と代謝障害を制御する。
Immunity. 2016; 45: 802-816
論文で見る
スコープス(437)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ヘーガンT.
プレンドラン B.
システム生物学はその約束を果たし、新規または改良型ワクチンの開発に貢献するか?システム生物学を通してデータから理解へ。
Cold Spring Harb. Perspect. Biol. 2018; 10: 028894
論文で見る
スコープス (26)
クロス
グーグル奨学生
ヘンリー・デュナン C.J.
レオン P.E.
ホアン M.
チョイ A.
クロミコバ V.
ホー I.Y.
タン G.S.
クルーズ J.
ハーシュ A.
鄭N.Y.
他
中和および非中和ヒトH7N9インフルエンザワクチン誘導モノクローナル抗体による防御。
Cell Host Microbe. 2016; 19: 800-813
論文で見る
スコープス (204)
PubMed
要旨
全文
全文PDF
グーグル奨学生
イリザリーR.A.
ホッブスB.
コリン F.
ビーザー・バークレイ Y.D.
アントネリス K.J.
シェルフ U.
スピードT.P.
高密度オリゴヌクレオチドアレイプローブレベルデータの探索、正規化、要約。
Biostatistics. 2003; 4: 249-264
論文で見る
筑波大学
PubMed
クロス
グーグル奨学生
ジャカール P.
高山植物の分布。
New Phytol. 1912; 11: 37-50
記事で見る
Scopus (3028)
クロスリファレンス
グーグル奨学生
川又陽子
藤井理恵子
細谷真一
原田宗男
吉田秀樹
三輪真理子
福住慎太郎
羽端康成
伊藤貴志
新谷祐子
他.
胆汁酸に応答するGタンパク質共役型受容体.
J. Biol. Chem. 2003; 278: 9435-9440
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
キーニー K.M.
ユリスト-ドウチュS.
アリエッタM.C.
フィンレイB.B.
ヒトの微生物叢とその後の疾患に対する抗生物質の影響。
Annu. Rev. Microbiol. 2014; 68: 217-235
論文で見る
スコープス (206)
PubMed
クロス
グーグル奨学生
クラーナ S.
ヴェルマ S.
ヴェルマN.
クレバー C.J.
カーター D.M.
マニシェウィッツ J.
キング L.R.
ロス T.M.
ゴールディング H.
適切に折り畳まれた細菌発現H1N1ヘマグルチニン球状頭部およびエクトドメインワクチンは、H1N1パンデミックインフルエンザウイルスからフェレットを保護する。
PLoS ONE. 2010; 5: e11548
論文で見る
スコープス (81)
PubMed
クロス
グーグル奨学生
クラーナ S.
ヴェルマN.
ユーデルJ.W.
ヒルベルト A.K.
カステッリーノ F.
ラッタンジ M.
デル・ジュディチェG.
ラプオリ R.
ゴールディング H.
MF59アジュバントは、パンデミックインフルエンザワクチンに対する抗体媒介免疫応答の多様性と親和性を高める。
Sci. Transl. Med. 2011; 3: 85ra48
論文で見る
スコープス (292)
PubMed
クロス
グーグル奨学生
クラーナ S.
ハーン M.
コイル E.M.
キング L.R.
リン T.L.
トレオナー J.
サント A.
ゴールディング H.
反復ワクチン接種により、ヒトにおける異なるインフルエンザワクチンプラットフォーム間での抗体親和性の成熟が抑制される。
Nat. Commun. 2019; 10: 3338
論文で見る
スコープス (54)
PubMed
クロス
グーグル奨学生
クラット N.R.
チェウ R.
ビルセ K.
ゼビン A.S.
ペルナー M.
ノエル・ロマスL.
グローブラー A.
ウェストマコットG.
謝 I.Y.
バトラーJ.
他。
膣内細菌がアフリカ人女性におけるHIVテノホビル殺微生物剤の有効性を修飾する。
Science. 2017; 356: 938-945
論文で見る
スコープス (279)
PubMed
クロス
グーグル奨学生
コスティッチ A.D.
ザビエルR.J.
ゲバースD.
炎症性腸疾患におけるマイクロバイオーム:現状と今後の展望。
Gastroenterology. 2014; 146: 1489-1499
論文で見る
スコープス(1200)
PubMed
要旨
全文
全文PDF
グーグル奨学生
Levine M.M.
開発途上国における経口ワクチンの免疫原性と有効性:コレラ生ワクチンからの教訓。
BMC Biol.
論文で見る
スコープス(214)
PubMed
クロスフィルム
グーグル奨学生
レヴィ M.
ブラッハーE.
エリナブE.
マイクロバイオーム、代謝産物と宿主免疫。
Curr. Opin. Microbiol. 2017; 35: 8-15
論文で見る
スコープス (291)
PubMed
クロス
グーグル奨学生
Li S.
Park Y.
ドゥライシンガム S.
Strobel F.H.
カーン N.
ソルトウ Q.A.
ジョーンズ D.P.
Pulendran B.
高スループットメタボロミクスからネットワーク活性を予測する。
PLoS Comput. Biol.
論文で見る
筑波大学
パブコメ
クロス
グーグル奨学生
Li S.
ルフェールN.
ドゥライシンガムS.
ロメロ-スタイナーS.
プレスネルS.
デイビス C.
シュミット D.S.
ジョンソン S.E.
ミルトン A.
ラジャムG.
他。
5種類のヒトワクチンのシステム生物学的研究から得られた抗体応答の分子シグネチャー。
Nat. Immunol. 2014; 15: 195-204
論文で見る
スコープス (475)
PubMed
クロス
グーグル奨学生
Li S.
サリバン N.L.
ルーファエル N.
ユー T.
バントン S.
マドゥール M.S.
マコウスランド M.
チウ C.
カニフ J.
ドベイ S.
ら。
ヒトにおけるワクチン接種に対する反応の代謝表現型。
Cell. 2017; 169: 862-877.e817
論文で見る
スコープス (174)
PubMed
要旨
全文
全文PDF
グーグル奨学生
リットマンD.R.
微生物叢はワクチンや病原体に対する防御免疫に影響するか?もしそうなら、有効な微生物叢に基づくワクチンの可能性はあるか?
Cold Spring Harb. Perspect. Biol. 2018; 10: a029355
論文で見る
スコープス (16)
PubMed
クロス
グーグル奨学生
牧島真一
Lu T.T.
Xie W.
ウィットフィールド G.K.
堂本裕一
エヴァンス R.M.
ハウスラー M.R.
マンゲルスドルフD.J.
腸内胆汁酸センサーとしてのビタミンD受容体。
Science. 2002; 296: 1313-1316
論文で見る
哺乳類
PubMed
クロス
グーグル奨学生
マティスV.
フリックE.
ゲファーズR.
ゲスリングE.
Haubrock M.
ヘール R.
ホルニッシャー K.
カラス D.
ケル A.E.
ケル・マルグリスO.V.
他。
TRANSFAC:転写制御、パターンからプロファイルへ。
核酸研究 2003; 31: 374-378
論文で見る
日本学術振興会特別研究員
PubMed
クロスリファレンス
グーグル奨学生
Mitre E.
スシ A.
クロップL.E.
シュワルツ D.J.
ゴーマン G.H.
Nylund C.M.
乳児期の酸抑制薬および抗生物質の使用と幼児期のアレルギー疾患との関連。
JAMA Pediatr. 2018; 172: e180315
論文で見る
スコープス(168)
PubMed
Crossref
グーグル奨学生
モディ S.R.
コリンズJ.J.
レルマンD.A.
抗生物質と腸内細菌叢。
J. Clin. Invest. 2014; 124: 4212-4218
論文で見る
スコープス (439)
PubMed
クロス
グーグル奨学生
中谷博之
Wrammert J.
リーE.K.
ラシオッピL.
マリー-クンツェS.
ヘイニング W.N.
ミーンズ A.R.
カストリ・S.P.
カーン N.
リーG.M.
他
ヒトにおける季節性インフルエンザワクチン接種のシステム生物学。
Nat. Immunol. 2011; 12: 786-795
論文で見る
スコープス (620)
PubMed
クロス
グーグル奨学生
中谷博之
ヘーガンT.
ドゥライシンガム S.S.
リー E.K.
クウィッサ M.
ルーファエル N.
フラスカ D.
ゲルステン M.
メータ A.K.
ガウジューR.
他
複数年にわたる多様な集団におけるインフルエンザワクチン接種に対する免疫のシステム解析により、分子シグネチャーの共有が明らかになった。
Immunity. 2015; 43: 1186-1198
論文で見る
スコープス (203)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ニコルソンJ.K.
ホームズE.
キンロスJ.
バーセリンR.
ギブソン G.
ジャ W.
ペターソンS.
宿主-腸内細菌叢代謝相互作用。
Science. 2012; 336: 1262-1267
論文で見る
スコパス(3146)
PubMed
クロス
グーグル奨学生
オー J.Z.
ラヴィンドラン R.
シャサイングB.
カルバーリョ F.A.
マドゥール M.S.
バウワー M.
ハキンプール P.
ギル K.P.
ナカヤ H.I.
ヤロヴィンスキー F.
他
季節性インフルエンザワクチン接種に対する抗体応答には、TLR5を介した腸内細菌叢の感知が必要である。
Immunity. 2014; 41: 478-492
論文で見る
スコープス (380)
PubMed
要旨
全文
全文PDF
グーグル奨学生
パブリディスP.
パウエル N.
ヴィンセント R.P.
エーリッヒD.
ビャルナソン I.
ヘイB.
システマティックレビュー:胆汁酸と腸の炎症-粘膜防御の管腔アグレッサーまたはレギュレータ?
アリメント。薬理学。Ther. 2015; 42: 802-817
論文で見る
スコープス (95)
PubMed
クロス
グーグル奨学生
ペイ L.
カストリロ A.
トントノズP.
オーファン核内受容体Nur77によるマクロファージ炎症遺伝子発現制御。
Mol. Endocrinol. 2006; 20: 786-794
論文で見る
スコープス (169)
PubMed
クロス
グーグル奨学生
Pulendran B.
システムワクシノロジー:ワクチンで人類の多様な免疫システムを探る。
Proc. Natl. Sci. USA. 2014; 111: 12300-12306
論文で見る
スコープス (124)
PubMed
クロス
グーグル奨学生
プレンドランB.
リー・S.
中谷博之
システムワクチン学。
Immunity. 2010; 33: 516-529
論文で見る
スコープス (299)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
ランペリ S.
カンデラ M.
トゥローニ S.
ビアジ E.
コッリーノ S.
フランチェスキ C.
オトゥール P.W.
ブリギディP.
極端な加齢における腸内細菌叢の機能的メタゲノム解析。
Aging (Albany N.Y.). 2013; 5: 902-912
論文で見る
スコープス(229)
PubMed
クロス
グーグル奨学生
Rappuoli R.
シエナ E.
フィンコ O.
システム生物学はその約束を果たし、新規あるいは改良型ワクチンの開発に貢献するか?ワクチンの自然免疫と適応免疫に関するシステム生物学の見解。
Cold Spring Harb. Perspect. Biol. 2018; 10: a029256
論文で見る
スコープス (7)
PubMed
クロス
グーグル奨学生
リドロンJ.M.
カン D.J.
ハイレモンP.B.
ヒト腸内細菌による胆汁酸塩の生体内変換。
J. 脂質研究 2006; 47: 241-259
論文で見る
(1869年)
PubMed
クロス
グーグル奨学生
リドロンJ.M.
カン D.J.
ハイレモンP.B.
バジャージJ.S.
胆汁酸と腸内マイクロバイオーム。
Curr. 意見。Gastroenterol. 2014; 30: 332-338
論文で見る
スコープス (814)
PubMed
クロス
グーグル奨学生
ルティ B.
ル・シャトリエE.
デローザL.
ドゥオン C.P.M.
Alou M.T.
ダイエール R.
フルキガー A.
メッサウデネ M.
ラウバー C.
ロベルティ M.P.
他
上皮性腫瘍に対するPD-1ベースの免疫療法の有効性に腸内細菌叢が影響する。
Science. 2018; 359: 91-97
論文で見る
スコープス (3070)
PubMed
クロス
グーグル奨学生
サラザールN.
バルデス-バレラL.
ゴンサレスS.
グエモンデM.
デ-ロス-レイエス-ガビランCG。
高齢者の栄養と腸内細菌叢。
腸内微生物。2017; 8: 82-97
論文で見る
スコープス (158)
PubMed
クロス
グーグル奨学生
サベージ D.C.
消化管の微生物生態学。
Annu. Rev. Microbiol. 1977; 31: 107-133
論文で見る
スコパス (1717)
PubMed
クロス
グーグル奨学生
Scher J.U.
シェスナックA.
ロングマンR.S.
セガタ N.
ウベダ C.
ビエルスキ C.
ロストロン T.
セルンドロ V.
パマー E.G.
エイブラムソンS.B.
et al.
腸内プレボテラ・コプリの増殖は、関節炎感受性の増強と相関する。
eLife. 2013; 2: e01202
論文で見る
PubMed
クロスレフ
グーグル奨学生
シュロス P.D.
ウェストコット S.L.
リャビンT.
ホール J.R.
ハルトマン M.
ホリスター E.B.
レスニウスキー R.A.
オークリーB.B.
パークス D.H.
ロビンソンC.J.
他
微生物群集の記述と比較のための、オープンソース、プラットフォーム非依存、コミュニティ支援型ソフトウェア。
Appl. Microbiol. 2009; 75: 7537-7541
論文で見る
筑波大学
PubMed
クロス
グーグル奨学生
シュミット N.
上野博之
血液Tfh細胞には色がある。
Immunity. 2013; 39: 629-630
論文で見る
スコープス (53)
パブコメ
概要
全文
全文PDF
グーグル奨学生
シャー Y.M.
マー X.
森村和彦
キム I.
ゴンザレス F.J.
プレグナンX受容体の活性化は、NF-κB標的遺伝子の発現抑制を介して黄砂誘発炎症性腸疾患を改善する。
Am. J. Physiol. Gastrointest. Liver Physiol: G1114-G1122
論文で見る
スコープス(203)
PubMed
クロス
グーグル奨学生
シャノン P.
マーキエルA.
オジエO.
バリガ N.S.
ワン J.T.
ラメイジ D.
アミン N.
シュビコフスキーB.
Ideker T.
Cytoscape: 生体分子相互作用ネットワークの統合モデルのためのソフトウェア環境。
ゲノム研究 2003; 13: 2498-2504
論文で見る
筑波大学
PubMed
クロスフィルム
グーグル奨学生
シンクレア C.
ボマカンティG.
ガルディナッシL.
ローバーマンJ.
ジョンソン M.J.
ハキンプール P.
ヘーガン T.
ベニテス L.
トドール A.
マキアD.
他。
mTORは肺におけるAPCの代謝適応を制御し、アレルギー性炎症の結果を制御する。
Science. 2017; 357: 1014-1021
論文で見る
スコープス (80)
PubMed
クロス
グーグル奨学生
ソネンブルグJ.L.
Bäckhed F.
ヒトの代謝のモデレータとして食事-微生物叢相互作用。
Nature. 2016; 535: 56-64
論文で見る
スコパス (1345)
PubMed
クロス
グーグル奨学生
スタウディンガーJ.L.
グッドウィンB.
ジョーンズ S.A.
ホーキンス-ブラウンD.
マッケンジー K.I.
ラトゥール A.
リュー Y.
クラッセン C.D.
ブラウン K.K.
ラインハルトJ.
他
核内受容体PXRは、肝毒性から保護するリトコール酸センサーである。
Proc. Natl. Acad. Sci. USA. 2001; 98: 3369-3374
論文で見る
(株)スコープス (1176)
PubMed
クロス
グーグル奨学生
シュタインホフU.
群衆は誰がコントロールしているのか?腸内細菌叢についての新しい知見と古い質問。
Immunol. Lett.
論文で見る
スコパス (54)
PubMed
クロス
グーグル奨学生
スブラマニアンA.
タマヨ P.
ムータ V.K.
ムカルジー S.
イベートB.L.
ジレット M.A.
パウロビッチ A.
ポメロイ S.L.
ゴルブ T.R.
ランダー E.S.
メシロフ J.P.
遺伝子セット濃縮解析:ゲノムワイド発現プロファイルを解釈するための知識ベースのアプローチ。
Proc. Natl. Sci. USA. 2005; 102: 15545-15550
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
タバニャナム S.
フレミングJ.
ブロムリーA.
シールズ M.D.
カードウェルC.R.
帝王切開と小児喘息との関連についてのメタアナリシス。
Clin. Exp. Allergy. 2008; 38: 629-633
論文で見る
スコープス (506)
PubMed
クロス
グーグル奨学生
テバランジャン N.
プクタA.
シュルツC.
ナイドゥー A.
Szamosi J.C.
ヴェルショール C.P.
ルーコフ D.
シェンク L.P.
ジュリー J.
フォーリー K.P.
他
加齢に伴う微生物性ディスバイオーシスは、腸管透過性、全身性炎症、マクロファージ機能不全を促進する。
Cell Host Microbe. 2017; 21: 455-466.e4
論文で見る
スコープス (620)
PubMed
要旨
全文
全文PDF
グーグル奨学生
トラークV.A.
ウォルトマンL.
ヴァン・エックN.J.
ルーヴァンからライデンへ:つながりのあるコミュニティを保証する。
Sci. Rep. 2019; 9: 5233
論文で見る
スコープス (1267)
PubMed
クロス
グーグル奨学生
トレマローリV.
Bäckhed F。
腸内細菌叢と宿主代謝の間の機能的相互作用。
Nature. 2012; 489: 242-249
論文で見る
スコパス(3143)
PubMed
クロス
グーグル奨学生
ヴォルカー T.
ワン・エイチ
アイリッシュ M.
Wang J.
ハリソン S.
ロイシェル S.
ベネット P.
Meng M.
規制バイオアナリシスのための6種類の胆汁酸のメソッド開発とバリデーション:選択性と感度の向上。
Bioanalysis. 2013; 5: 1229-1248
論文で見る
胆汁酸
PubMed
クロス
グーグル奨学生
ウランマートJ.
スミス K.
ミラーJ.
ラングレー W.A.
コッコ K.
ラーセン C.
鄭 N.Y.
メイズ I.
ガーマン L.
ヘルムズC.
他。
インフルエンザウイルスに対する高親和性ヒトモノクローナル抗体の迅速クローニング。
Nature. 2008; 453: 667-671
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
ヤツネンコ T.
レイ F.E.
マナリー・M.J.
トレハン I.
ドミンゲス・ベロ M.G.
コントレラス M.
マグリス M.
イダルゴ G.
バルダッサーノ R.N.
アノヒンA.P.
他。
ヒトの腸内細菌叢は年齢と地理に関係なく見ることができる。
Nature. 2012; 486: 222-227
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
Yu T.
パーク Y.
ジョンソン J.M.
ジョーンズ D.P.
apLCMS--高分解能LC/MSデータの適応的処理。
Bioinformatics. 2009; 25: 1930-1936
論文で見る
スコープス (253)
PubMed
クロスリファレンス
グーグル奨学生
Zitvogel L.
マー Y.
ラウルD.
クローマー G.
ガジェフスキーT.F.
癌免疫療法におけるマイクロバイオーム: 診断ツールと治療戦略。
Science. 2018; 359: 1366-1370
論文で見る
スコープス (433)
PubMed
クロス
グーグル奨学生
論文情報
出版履歴
掲載 2019年9月5日
受理済み 2019年8月6日
改訂版受理 2019年6月21日
受理 2018年8月28日
身分証明書
DOI: https://doi.org/10.1016/j.cell.2019.08.010
著作権
© 2019 Elsevier Inc.
ユーザーライセンス
Elsevierユーザーライセンス|情報アイコンの再利用方法
サイエンスダイレクト
ScienceDirectでこの記事にアクセスする
図
図サムネイルfx1
グラフィカルアブストラクト
図サムネイルgr1
図1研究の概要と健康成人の腸内細菌叢に対する抗生物質使用の影響
図サムネイルfigs1
図S1図1に関連するフェーズ2における健康成人の腸内細菌叢に対する抗生物質使用の影響
図サムネイルgr2
図2抗生物質投与がTIVに対する体液性反応に及ぼす影響
図サムネイルfigs2
図S2TIVに対する体液性反応の追加測定(図2に関連
サムネイル図3
図S3図2に関連したTIVに対する適応的細胞応答
サムネイルgr3
図3対照および抗生物質治療被験者のTIVに対する転写反応
図サムネイルfigs4
図S4図3、5および6に関連した、第1相および第2相における転写および代謝反応の比較
図サムネイルgr4
図4抗生物質投与に対する転写および細胞反応
図サムネイルgr5
図5抗生物質投与とインフルエンザワクチン接種が血液メタボロームに及ぼす影響
図サムネイルgr6
図6二次胆汁酸の攪乱は抗生物質投与被験者におけるNLRP3インフラマソームシグナル伝達の上昇と関連している
図のサムネイルfigs5
図S5 図7に関連するMMRN構築のワークフロー
図サムネイルgr7
図7MMRN解析から、炎症シグナル伝達とH1N1特異的IgG1応答の制御における腸内細菌叢の異なる機能が示唆される
関連記事
広告
ライフ&メディカルサイエンスジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノム学
細胞宿主と微生物
細胞代謝
細胞レポート
セルレポーツ医学
細胞幹細胞
細胞システム
カレントバイオロジー
発生細胞
免疫
医学
分子細胞
ニューロン
構造
アメリカン・ジャーナル・オブ・ヒューマン・ジェネティクス (パートナー)
生物物理学雑誌 (パートナー)
生物物理レポート (パートナー)
Human Genetics and Genomics Advances (パートナー)
分子植物 (パートナー)
分子療法 (パートナー)
分子治療法と臨床開発 (パートナー)
分子療法核酸 (パートナー)
分子療法腫瘍学 (パートナー)
Plant Communications (パートナー)
幹細胞レポート (パートナー)
生化学の動向
癌の動向
細胞生物学の動向
生態学と進化の動向
内分泌学・代謝学の動向
遺伝学の動向
免疫学の動向
微生物学の動向
分子医学の動向
神経科学の動向
寄生虫学の動向
薬理学の動向
植物科学の動向
物理科学・工学ジャーナル
セルレポート 物理科学
化学
化学触媒
デバイス
ジュール
物質
化学の動向
学際ジャーナル
セルリポーツ・メソッド
セルリポーツ・サステイナビリティ
Heliyon
アイサイエンス
ワン・アース
パターン
STARプロトコル
ネクサス(パートナー)
イノベーション(パートナー)
バイオテクノロジーの動向
認知科学の動向
著者
論文投稿
複数ジャーナル投稿
STARメソッド
プレビュー - プレプリント
査読者
査読者向け情報
ニュース&イベント
ニュースルーム
細胞シンポジウム
コンソーシアムハブ
ウェビナー
ラボリンク
マルチメディア
セルプレスポッドキャスト
セルプレスビデオ
カラーリングとコミック
フィギュア360
セル画ショー
研究篇
セルプレスについて
セルプレスについて
オープンアクセス
COVIDハブ
持続可能性
インクルージョンと多様性
コンタクト
お問い合わせ
ヘルプ&サポート
採用情報
セルプレス採用情報
サイエンティフィックジョブボード
アクセス
登録する
請求
今すぐ読む
司書に推薦する
出版アラート
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
Nucleusコレクション
スナップショット・アーカイブ
インフォメーション
広告主の皆様へ
リクルーターの方へ
図書館員の方へ
プライバシーポリシー
ご利用条件
アクセシビリティ
本サイトのコンテンツは、あらゆる分野の医療従事者および研究者を対象としています。
当サイトでは、サービスの提供・向上やコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を更新するには、このサイトのクッキー設定をご覧ください。
このサイトのすべてのコンテンツ: 著作権 © 2023 Elsevier Inc.
テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビア社に帰属します。
すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条件が適用されます。
プライバシーポリシー 利用規約 アクセシビリティ ヘルプ&サポート お問い合わせ
RELX
この記事が気に入ったらサポートをしてみませんか?