
遺伝現象と関連する物質

★遺伝情報とその発現
遺伝子の本体はDNAである。
そして各遺伝子の情報はヌクレオチドの塩基配列として書き込まれている。
その為、遺伝子を利用する時にはこれを読み出し具現化する。
これを発現と呼ぶ。
多くの遺伝子は様々な機能を持つタンパク質の設計図であるため、発現に伴ってタンパク質の合成が起こる。
・ 核酸
DNAとRNAの事。ヌクレオチドのポリマー。
それぞれ4種類のヌクレオチドから構成されている。
DNAは塩基間の相補性を利用して二重らせん構造をとり、RNAは一本鎖で様々な立体構造をとることができる(リボザイムや tRNA 等)。
また、リン酸ジエステル結合を形成(化学反応に必要なエネルギーは高エネルギーリン酸結合から得る)することでヌクレオチド同士はポリマーになっていく。
さらに。DNAとRNAの合成に関しては大きな違いが2つある。
一つは使われるヌクレオチド、もう一つがプライマーである。
DNAの複製(合成)ではプライマーと呼ばれる短いヌクレオチド鎖が必要で、RNAの合成では必要ない。
・ 染色体とDNA
染色体は階層構造を持つ。
DNAはヒストンというタンパク質に巻き付いた状態(ヌクレオソームと呼ばれる)で存在する。さらにヌクレオソームは、繊維状の構造(クロマチン繊維)を形成し、折りたたまれて棒状の構造をとる。これが染色体である。
● 細胞周期におけるDNA量の変化
細胞が分裂を開始してから次の分裂を開始するまでの1サイクルを細胞周期という。これはG1期、S期、G2期、M期の4つのステージから成る。
S期にはDNAの複製が行われ細胞のDNA量は倍増する。そして、細胞分裂が終了すると2つに複製されたDNAがそれぞれ異なる細胞に完全に振り分けられる。従って、1つの細胞中のDNA量はまず倍増し、その後半減する。
● DNAの複製の仕組み
S期におけるDNA合成反応(複製)は、DNAの二重らせん構造から理論的に予測された。これは半保存的複製と呼ばれている。
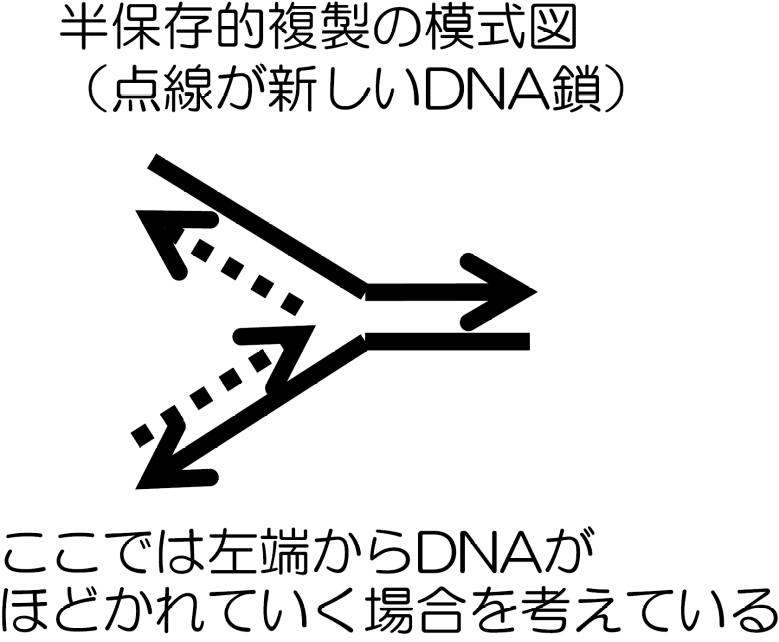
この反応では遺伝情報を正確にコピーするために塩基の相補性を利用する。つまり、DNAの二重らせんをほどいて半分だけ新しいDNA鎖を作る。
これがその仕組みの名前の由来である。
また、この反応には各種のデオキシリボヌクレオシド三リン酸(dNTP)やプライマー、DNAポリメラーゼ(DNA合成酵素)という材料や酵素が必要であり、化学反応に必要なエネルギーはdNTPによって供給される。
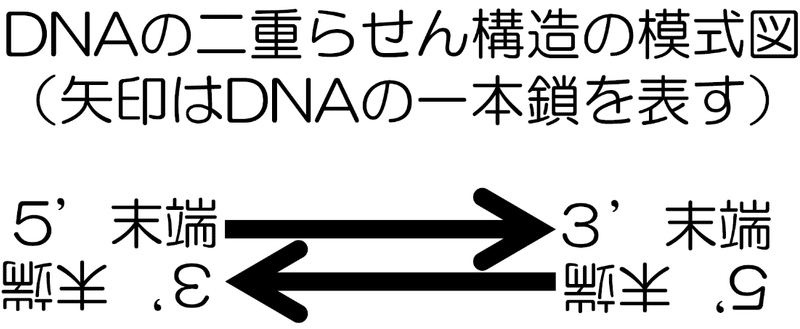
・ DNAの方向性
ヌクレオチドは上下に非対称な分子である(五角形の五炭糖が原因)。
さらに、DNAはそのヌクレオチドが同じ向きに規則的に結合してできる。
従ってDNAも非対称となる。これをDNAの方向性(極性)という。
また、この性質はDNAの末端が区別できる事とも関連している。
具体的には、リン酸が突き出ている末端はこれが結合している五炭糖の炭素の番号から5’ 末端と呼ばれ、もう一方のヒドロキシ基が突き出ている末端はそれが結合している五炭糖の炭素の番号から3’ 末端と呼ばれて区別される。
それに加えて、DNAの二重らせん構造は相補性の性質により方向性が反対の2本のDNA鎖が結びついた形をとる(逆平行二本鎖)。従って、二重らせん構造をとっている2本鎖DNAの端を見ると、5’ 末端と3’ 末端が結びついている形になっている。
・ 不連続なDNAの複製
DNAの方向性に関連してDNAポリメラーゼにも機能の非対称性が見られる。この酵素は5’ 末端から3’ 末端方向にしかDNAの合成ができない。しかし、二本鎖DNAを一定方向にほどいていく場合、その構造が逆平行の二本鎖であるため片方のDNA鎖ではどうしても 3’ 末端から 5’ 末端方向にDNAの合成が行われる必要がある。どのようにこの矛盾を解決するのだろうか?
*以下の模式図の注意点。
左から右方向にDNAをほどきながら複製を行う場合の話と図である!
左からDNAをほどきながら複製するので左から右向きに複製も進む。
2本鎖下側のDNAは 5’ 末端から 3’ 末端方向に複製が行えるので問題ない。
しかし、上側では2本鎖DNAをほどく方向と複製する方向が逆になってしまう。これは右から左にDNAをほどきながら複製する場合も同様で、結局上下どちらかで、ほどく方向と複製する方向が食い違ってしまう。
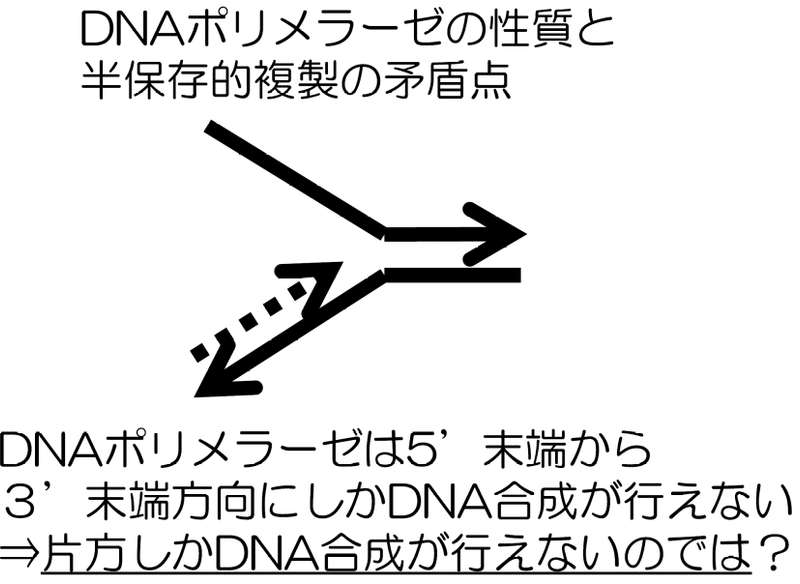
この問題の解決策は単純である。
DNAポリメラーゼをDNAをほどく方向に対して逆走させる。
つまり、DNAポリメラーゼをDNAをほどく向きと逆方向に働かせる。そのため、二重らせんをほどきながらの即座にDNAの合成はできず、いくらかほどかれてしまった後に、その都度短いDNAを合成していく。そして、それらをつなぎ合わせてひとつながりの連続したDNA鎖を作る。
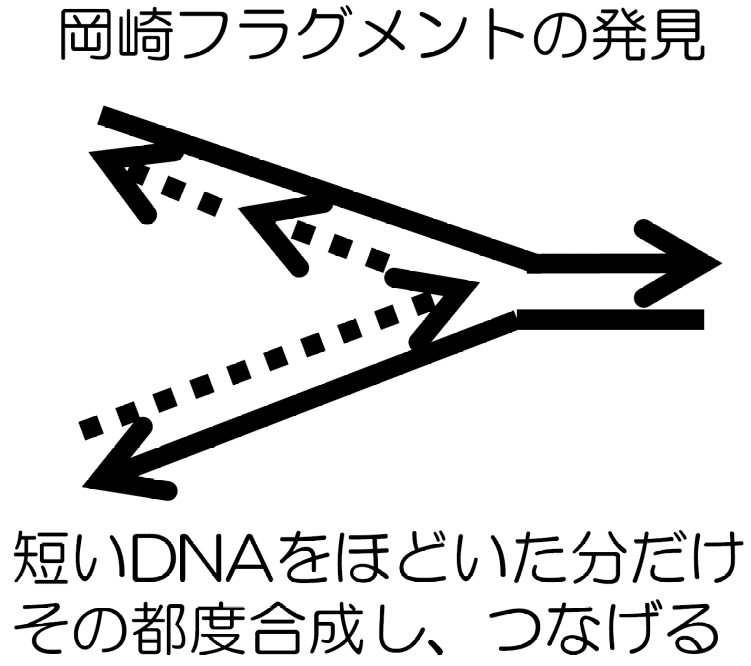
これは、広島出身の岡崎令治により発見された。ゆえに、この短いDNA断片を岡崎フラグメントと呼ぶ。岡崎フラグメントが結合して作られる新DNA鎖はDNA合成に時間がかかる(タイムラグがある)。そのためラギング鎖と呼ばれる。もう一方は合成が速く進みリードするのでリーディング鎖と呼ばれている。
このような仕組みは一見非合理に見える。しかし、おそらくミスを防ぐための方法とのトレードオフだと考えられる。DNA ポリメラーゼを 3’ 末端から 5’ 末端に動かすと結合させたヌクレオチドを切り離せるからだ。
すなわち、間違った時にそれを修正するための機能として、逆向きの反応を手に入れた代償なのだ。岡崎フラグメントをせっせと作り、つなげてラギング鎖にするコストよりも、DNAの複製を間違って修正できずに突然変異としてしまうリスクを下げた方が利益が大きいのだろう。
突然変異をできるだけ起こしたくないのである。
●半保存的複製の証明(メセルソンとスタール)
メセルソンとスタールは、窒素の同位体と密度勾配遠心法を利用して半保存的複製を証明した。
・ 密度勾配遠心法
密度勾配が形成された状態で行う遠心分離法。イメージは風船の浮き沈み。ヘリウムガスを入れた風船は、空気の比重より軽いため浮く。だが、空気より重いガスを入れた風船は沈む。では、空気を入れた風船はどうなるだろうか。原理的には浮きも沈みもしない(留まる)はずである。
これを利用したのが密度勾配遠心法であり、混合物を密度により分離する。まず、下に行くほど密度の高い溶液(密度勾配を持つ溶液)を用意する。そこに混合物を入れると、物質は沈む。だが、各物質は自身の密度が、溶液の密度と同じになるところで沈まなくなる。その結果、物質が密度勾配に応じて分けられる。物質が沈むのに時間がかかる為、適度な遠心力で沈ませる。
・ メセルソンとスタールの実験
⓪目的と方法
DNAの複製方法を知るために、新しいDNAと古いDNAを見分け比較したい。だが、違いが無いので見分けがつかず比較できない。だから、新しいDNAと古いDNAに違い(差)を作り出す。
①重いDNAの作成
15N(重窒素)のみを生物に与えると、DNAが重くなる(重いDNA)。
②軽いDNAの作成
14N(窒素)しか与えない条件に切り替えると、新しいDNAは軽くなる(軽いDNA)。
③密度勾配遠心法を利用した密度によるDNAの分離
DNAの重さ(密度)の変化、及び重さごとのDNAの比率を密度勾配遠心法で数世代に渡って観察する(DNAの沈む位置やその濃さで重さや量が判明)。
①から②の条件に切り替えることで密度は低下していくので、その様子を調べていく。
④一回だけのDNA複製後の観察結果
一回のDNA複製後、軽いDNAと重いDNAの中間の重さのDNAのみが生じた。
⑤二回DNA複製後の観察結果
二回目のDNA複製後は、軽いDNAと中間の重さのDNAが1対1の割合で生じた。
④、⑤のような観察パターンは半保存的複製でしか説明できない。
(半保存的複製の証明)
・DNAの複製のライセンス化(真核細胞)
複製は一回の分裂に対して一回だけ起こるように制御されている。
これをライセンス化と呼ぶ。簡単にいえば、複製期以外は複製開始点にポリメラーゼが結合できないようにブロックするという仕組みである。
この記事が気に入ったらサポートをしてみませんか?