骨の修復・再構築の生理学:流体剪断応力が注目されている!?

骨の生理学
骨は高度に特殊化された硬い組織であり、身体を構造的に支持し、筋肉付着部位を通る動きを可能にし、臓器を保護し、カルシウムと成長因子の貯蔵庫として機能します(Clarke、2008 )。骨は生涯を通じて絶えず再生し修復する力を持っています。骨リモデリングと呼ばれるこのプロセスには、さまざまな種類の細胞が関与しており、生体力学的負荷の変化に応じて開始されたり、古い微小損傷を受けた骨を機械的により強い新しい骨に置き換えたりすることができます (Kini and Nandeesh、2012 )。
骨の再構築
骨のリモデリングは、骨の強度とミネラルの恒常性を維持するために不可欠なプロセスです。再構築により、古く損傷した骨を修復し、外部負荷の変化に合わせて骨の構造を調整することができます。このプロセスでは、特殊な細胞、つまり石灰化した基質を除去する破骨細胞と新しい骨基質を堆積する骨芽細胞が連携して働きます。それらの連携は生化学経路を通じて厳密に制御されています ( Hadjidakis and Androulakis, 2006 )。たとえば、骨芽細胞による核因子カッパ-B リガンドの受容体活性化因子 (RANKL) の放出は、破骨細胞前駆体の表面にある RANK 受容体への結合を通じて破骨細胞の活性化を誘導します。このプロセスは、RANKL に競合的に結合するオステオプロテゲリン (OPG) によって阻害される可能性があります ( Boyce および Xing、2008 )。リモデリング サイクル は、4 つの連続したフェーズで構成されます ( Clarke、2008 )。
•活性化: ホルモンまたは物理的刺激により、単核前破骨細胞が循環から骨再構築部位に動員されます。骨表面に付着した後、細胞は多核破骨細胞に融合します。
•吸収: 破骨細胞は、有機および無機骨成分の吸収を開始します。これには 2 ~ 4 週間かかります。破骨細胞は、小柱骨に特徴的なハウシップ裂孔を形成し、皮質骨に切断円錐を形成します。これらの空洞が一定のサイズに達すると、破骨細胞のアポトーシスによって骨吸収が終了します ( Sikavitsas et al., 2001 )。
•反転: 吸収された表面は単核マクロファージ様細胞によって平滑化され、マトリックス沈着の準備が整います。
•形成: 骨芽細胞は、コラーゲンマトリックスを分泌し、その石灰化を制御することによって新しい骨を築きます。このプロセスを通じて、一部の骨芽細胞はマトリックス内に埋もれ、完全に石灰化したラクナ小管系 (LCS) に存在する骨細胞に分化します。4 ~ 6 か月後、この段階は完了し、骨芽細胞は骨を裏打ちする細胞に変わるか、アポトーシスに入ります。
皮質骨では、骨強度を維持するには年間 2 ~ 3% の再構築率で十分です。海綿骨は代謝回転率が高く、カルシウムとリンの代謝にとって骨のリモデリングが重要です。
骨細胞
骨細胞は、新しい骨組織を堆積する骨芽細胞、骨基質を破壊する破骨細胞、機械的負荷に応じて骨芽細胞と破骨細胞の活動を調整する骨細胞の間のバランスを維持することにより、骨リモデリング中に協調して連携します(Hadjidakis and Androulakis、2006 ;ボーンワルドとジョンソン、2008 年)。
骨芽細胞
骨芽細胞は、間葉幹細胞 (MSC) に由来する骨形成細胞です ( Caplan、1991 )。MSCは適切な刺激を受けると骨芽細胞に分化しますが、軟骨、筋肉、腱、脂肪細胞に変化することもあります( Caplan and Bruder, 2001 )。骨芽細胞の分化と成熟のプロセスは、機械的経路と生化学的経路の両方によって支配されます。例えば、Runt関連転写因子2(Runx2)は前骨芽細胞の発生に不可欠であり、オステオポンチン、I型コラーゲン、オステオカルシン、アルカリホスファターゼ(ALP)などの骨芽細胞特異的遺伝子を活性化する(Ducyら、1997年、Xuら) al.、2015)。成熟骨芽細胞の分化は、ホルモンまたは機械的に活性化される Wnt シグナル伝達経路によって制御されます ( Westendorf et al., 2004 )。
前骨芽細胞の形態は線維芽細胞に非常に似ています。ただし、後者は石化されたマトリックスを生成できません。成熟した骨芽細胞は、典型的には立方体の形状をしている( Franz-Odendaal et al., 2006 )。骨芽細胞は、独自の分泌機構により骨基質の合成と石灰化を直接調節します。骨吸収は、破骨細胞に作用するパラクリン因子を介して骨芽細胞によって間接的に制御されます。例えば、RANKLの受容体活性化因子の放出は、破骨細胞前駆体の表面上のRANK受容体への結合を通じて骨吸収を開始する( Boyce and Xing、2008 )。骨芽細胞の平均寿命は、数日から約100日の範囲です( Rosenberg et al., 2012 )。骨芽細胞は、寿命の終わりに、
(1) 新たに形成された骨基質に埋め込まれて骨細胞に分化する、
(2) 不活性な骨表面を保護する不活性な骨裏打ち細胞に変化する、
(3) アポトーシスを開始する、
のいずれかになります (Manolagas 、 2000年)。

骨細胞
骨細胞は、新たに沈着した骨基質内に捕捉された、最終分化した骨芽細胞である( Franz-Odendaal et al., 2006 )。骨芽細胞と骨細胞は起源は同じですが、形態と機能が大きく異なります。骨細胞形成中、すなわち骨芽細胞から骨細胞への分化中、細胞体サイズは減少し、細胞突起は初期骨細胞のマーカーであるE11/gp38によって制御されている石灰化マトリックスに向かって放射状に広がり始める(Schulze et al., 1999 )。移行後は、ALP、I 型コラーゲン、骨形成タンパク質 2 (BMP-2) の遺伝子発現が減少します。オステオカルシン、E11/gp38、スクレロスチン(Sost)、象牙質基質タンパク質1(DMP-1)などの他のタンパク質も上方制御または導入されています(Mullen et al.、2013)。
骨細胞形成を調節する合図についてはほとんど知識がありません ( Dallas et al., 2013 )。石灰化した骨組織と比較して柔らかい、沈着した類骨の機械的特性が分化を導く可能性がある( Mullen et al., 2013 )。さらに、類骨の石灰化および低酸素状態も骨細胞形成の原動力である可能性がある( Irie et al., 2008 ; Prideaux et al., 2012 )。
骨細胞はもはや「骨の受動的なプレースホルダー」ではなく、非常に異なる機能を持つ細胞として見なされているため、骨細胞の研究は近年関心を集めています( Bonewald、2011 )。骨細胞は骨の中で最も豊富な細胞種(全骨細胞の 90 ~ 95%)であり、シグナル因子を放出することで機械的負荷に応答すると考えられています。これらの因子を通じて、それらは破骨細胞および骨芽細胞の活性を調節することによって骨のリモデリングを調整する( Knothe Tail et al., 2004 )。
破骨細胞
破骨細胞は、酸および溶解酵素を分泌することによって石灰化した骨基質を再吸収できる特殊な細胞です。これらは、骨髄に存在する単核前駆細胞に由来する多核細胞です ( Boyle et al., 2003 )。それらの分化 (破骨細胞形成) は、隣接する間質細胞や骨芽細胞によって産生される RANKL やマクロファージ コロニー刺激因子 (M-CSF) などのサイトカインによって制御されます。破骨細胞の分化は、高い親和性でRANKLに結合し、RANK受容体への結合を妨げるOPGによって阻害され得る( Suda et al., 1999 )。
骨細胞外マトリックス
骨の細胞外マトリックス (ECM) は、50 ~ 70% の無機ミネラル、20 ~ 40% の有機材料、3% 未満の脂質、および水からなる複合材料です (Clarke、2008 )。正確な組成は、年齢、骨の部位、性別、骨粗鬆症などの病状などの要因によって異なります ( Boskey、2013 )。
骨のミネラル部分はヒドロキシアパタイトによく似ており、骨に機械的剛性と耐荷重強度を与えます ( Boskey、2007 )。この相は、ナトリウム、マグネシウム、クエン酸塩、フッ化物などの不純物も含むカルシウムとリン酸塩の結晶複合体として最もよく説明できます ( Khan et al., 2013 )。骨の弾性と柔軟性は、コラーゲンやフィブロネクチンなどの構造タンパク質を含む有機成分によって提供されます ( Nair et al., 2013 )。有機相は、マトリックスの組織化とミネラルの沈着を制御する重要な機能を果たす他の非コラーゲン性マトリックスタンパク質からも構成されています ( Young, 2003 ; Boskey, 2013 )。例えば、石灰化は小さなCa 2+結合タンパク質オステオカルシンによって制御されている可能性があります。機械的伝達は、細胞表面のインテグリンに結合できるオステオポンチンやオステオネクチンなどの糖タンパク質によって促進されます。オステオポンチンは、破骨細胞の骨表面への付着も可能にします ( Gundberg、2003 )。少量の脂質は細胞シグナル伝達とイオンの流れにとって重要です ( Clarke、2008 )。

コラーゲン集合体
コラーゲン原線維の集合は、細胞内および細胞外のステップを含む複雑なプロセスです。コラーゲンは、これらの分子が細胞の外で長い繊維に組み立てられる前に、まず細胞内空間で前駆体分子(プロコラーゲン)として合成されます。
コラーゲンの形成は、骨芽細胞や線維芽細胞などのコラーゲン産生細胞の核で始まります。核内では、デオキシリボ核酸 (DNA) の特定のセグメントがメッセンジャーリボ核酸 (mRNA) に転写されます。mRNA が核から細胞質に移動した後、プレプロコラーゲンとして知られるポリペプチド鎖に翻訳されます。各鎖の長さは約 300 nm、直径は 1.5 nm です。それらは、Gly-Y-Z の複数のトリプレット配列からなる厳密なパターンによって特徴付けられます。後でこれらの鎖が適切に折りたたまれるように、グリシン残基 (Gly) が 3 番目ごとの位置に存在する必要があります。Y と Z は任意のアミノ酸でありえますが、一般的にはプロリンとヒドロキシプロリンです ( van der Rest and Garrone、1991 )。各鎖は、両側のいくつかの特徴的なアミノプロペプチドとカルボキシプロペプチドによって終結します。これらの末端プロペプチドは、細胞内の長いコラーゲン線維の自己集合を防ぐのに不可欠です。
次に、小胞体 (ER) でプロリンおよびリジン残基がヒドロキシル化され、後のペプチド鎖の架橋を促進します。この酵素ステップには、補因子としてアスコルビン酸 (ビタミン C) が必要です。アスコルビン酸が不足すると、緩いコラーゲン三重らせんが形成されるか、コラーゲン合成が完全に妨げられ、壊血病などの疾患が引き起こされます ( Canty and Kadler、2005 )。3 つの修飾ペプチド鎖は三重らせんを形成し、ジスルフィド結合によってさらに安定化されます。I 型コラーゲンの場合、2 つのα 1 鎖と 1 つのα 2 鎖が集合して、プロコラーゲンと呼ばれる三重らせんを形成します。
タンパク質はらせん構造を形成した後、小胞体からゴルジ体に移動し、そこで分泌小胞に詰め込まれます。これらの担体はサイズと形態が異なり、長さ約 500 nm の液胞 ( Leblond、1989 )、またはより大きな管状嚢状構造 ( Polishchuk et al.、2000 )として説明されています。小胞は微小管に沿って原形質膜に向かって移動し、そこで細胞外空間にプロコラーゲンを放出します。
プロコラーゲンが細胞から放出されると、コラーゲン線維が細胞膜上に直接形成され始めます。この近接性により、細胞は線維形成を直接制御できる可能性があり、場合によっては腱や靱帯の平行な束や骨の絡み合い織りなどの長距離集合体の形成さえも制御できる可能性がある(Kadler et al., 2008 )。
フィブロネクチンおよびコラーゲン結合α 2 β 1 インテグリンなどの特定の細胞表面インテグリンは、線維状コラーゲンの組織化および沈着に必須であることが判明している(McDonald et al., 1982 ; Li et al., 2003)。ウェンストラップら(2004)。また、 in vivo でのコラーゲン I 線維の線維形成の誘導には、少量の V 型コラーゲンが必要であることも発見しました。
コラーゲン線維は、特定の酵素がトロポコラーゲンと呼ばれるプロコラーゲンから末端プロペプチドを除去した後にのみ形成できます( Prockop et al., 1998 )。トロポコラーゲン単位は自発的に集合してコラーゲン線維になります。数百個のトロポコラーゲン分子が特徴的な「四分の一千鳥」配列で並ぶため、複合繊維は電子顕微鏡で縞模様のように見えます。縞模様は、分子の縦方向の千鳥配置によって生じ、ある分子の終わりと次の分子の始まりの間に、トロポコラーゲンの長さ (67 nm) のおよそ 4 分の 1 の大きさの「穴」が残ります (Scott、1995 )。線維は共有結合の形成によってさらにサポートされます。酵素リシルオキシダーゼは、リジン上のヒドロキシル基およびヒドロキシルリジンをアルデヒド基に変換することによって結合の形成を触媒する( Kagan and Li、2003 )。その結果、側方および端から端までの融合の後、線維は直径が最大 10 倍、長さが劇的に増加します ( Birk et al., 1995 )。
コラーゲンの方向性
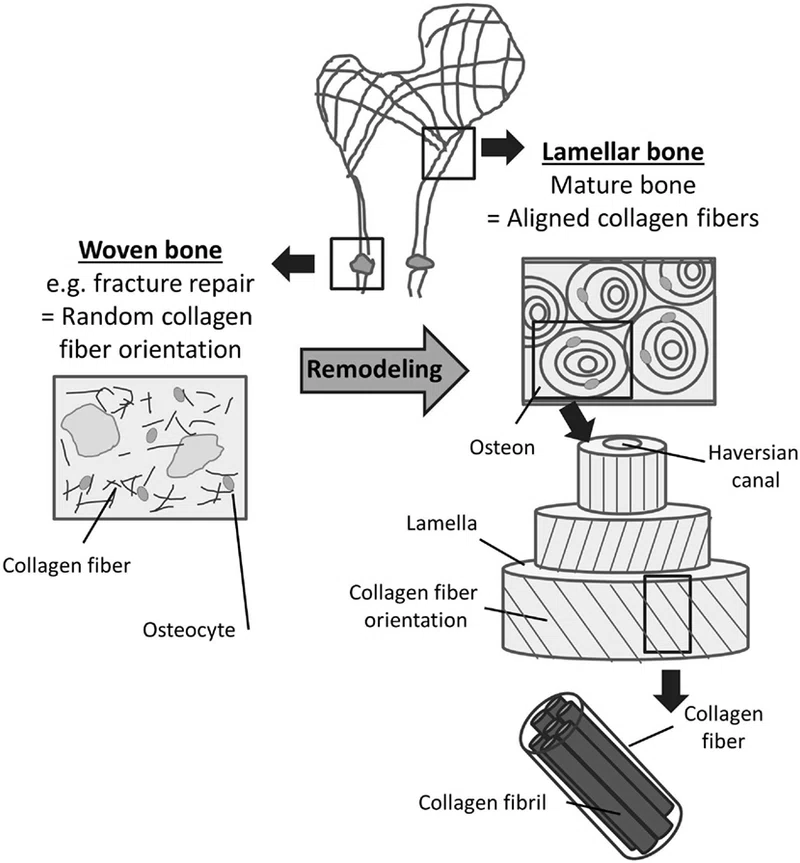
2 種類の骨は、骨基質内のコラーゲン線維の配向に基づいて区別できます:
(1) ランダムな配向のコラーゲン線維からなる織った骨、
(2) 高度に配向したコラーゲン線維によって特徴付けられる層状骨 (Kini および Nandeesh 、 2012年)。
層状骨のコラーゲン線維は平行な線維の配列で配置されており、順番に方向を変えて「ねじれた合板のような」構造を形成しています(Weiner et al.、1997)。コラーゲン配列の交互の配向により、編まれた骨と比較して層状骨の強度が大幅に高くなります ( Clarke、2008 )。この合板構造は、緻密な骨の主要な構成要素である円筒形の骨骨に見られます。オステオンは、血液と神経の供給を含むハバース管の周りに異なる方向に配置されたいくつかの同心円状の薄板で構成されています。オステオンの長軸は通常、骨の長軸と平行であり、支配的なコラーゲン線維の配向は一般に荷重の方向に従います ( Martin and Boardman, 1993 ; Hert et al., 1994 ; Seattle et al., 2008 )。
さらに、縦方向のコラーゲン線維は主に引張荷重を支える領域に存在し、圧縮荷重がかかる領域は横方向の線維で構成されます( Riggs et al., 1993 ; Martin et al., 1996 )。
コラーゲン線維の空間的配向に加えて、骨の機械的特性は、総骨量および骨ミネラルの材料特性を含む他のいくつかの要因によっても決定されます (Viguet -Carrin et al., 2006 )。後者は骨の剛性と降伏強度に関してより重要ですが、コラーゲンネットワークは骨の極限強度や靱性を含む降伏後の特性に大きな影響を与えるようです(Garnero、2015 )。線維配向が骨の強度に依存することは十分に受け入れられていますが、コラーゲンの配向が骨の機械的特性に与える影響を正確に推定することは依然として困難です (Viguet-Carrin et al., 2006 )。
機械的に弱い織られた骨は、骨基質が存在しない状況、つまり胎児および新生児の骨形成中、または骨折や骨切り後の骨修復の初期段階で生成されます。このような条件下では、間葉系骨芽細胞はコラーゲン線維をあらゆる方向に急速かつランダムに分泌します ( Shapiro, 1988 , 2008 )。しかしながら、上記のリモデリングプロセスの結果として形成される健康な成人の骨のほとんどは、高度に配向した層状骨で構成されています。平行なコラーゲン線維の生成には、骨生成細胞の集合的で組織的な作用が必要であると考えられています ( Kerschnitzki et al., 2011 )。
機械的刺激は、3D マトリックス配置を調整する別のトリガーである可能性があります。例えば、骨芽細胞は、おそらく骨芽細胞に加えられるひずみを最小限に抑え、より組織化されたコラーゲン束を分泌するために、周期的な伸張下で集合的に方向を変える( Matgaki et al., 2013 )。コラーゲン線維の整列に対する機械的力の影響は、線維芽細胞が石灰化されていないコラーゲンマトリックスを沈着させる軟組織において広く研究されています。たとえば、間質液の流れは、線維芽細胞を介したコラーゲンの整列に対する機械的刺激として作用する可能性があります。
骨においては、骨芽細胞によるコラーゲン線維の整列に関与する機構はまだ十分に理解されていない(matsugaki et al., 2013)。軟組織では、線維芽細胞によって発揮される牽引力がECMのリモデリングの主な推進力であると考えられています(Harris et al., 1981 ; Feng et al., 2014)。例えば、筋線維芽細胞の収縮とこれらの力のコラーゲンネットワークへの伝達は、創傷治癒における重要なステップであると考えられている( Tomasek et al., 2002 )。骨芽細胞によって加えられる牽引力は、線維芽細胞で観察されたのと同様の方法で線維を配向させることができる可能性がある( Curtze et al., 2004 )。Poellmannらは、牽引力顕微鏡を使用して、(2015)は、分化したコラーゲン産生骨芽細胞が、まだコラーゲン分泌に関与していない骨髄内に見られる前骨芽細胞と比較して、より高い牽引力を発揮することを実証しました。
さらに、微速度撮影画像は、細胞の運動性と組み合わされた牽引力により、骨芽細胞が線維状物質の「パケット」を移動させてECMを積極的に再構成できることを示唆しています( Dallas, 2006 )。骨芽細胞によって誘発される牽引力は、類骨層内に歪みを生じさせ、歪みのないコラーゲン線維の分解を通じてコラーゲン線維の整列を開始する可能性もあります(Flynn et al., 2010 ; Heck et al., 2015)。さらに、フィブロネクチン、インテグリン、およびコラーゲン V などの他のコラーゲン間の相互作用も、生体内でのコラーゲン線維配向プロセスに役割を果たしている可能性があります( Kadler et al., 2008 )。
石灰化
骨内の有機コラーゲンマトリックスは、ミネラル相によって強化されています。有機コラーゲンマトリックスの沈着と同様に、石化も細胞によって制御されるプロセスです。たとえば、骨芽細胞は選択された非ランダムな部位で石灰化を開始し、ECM へのイオン流を調節します ( Boskey、2007 )。
石灰化は 2 つの段階で起こります。まず、ハイドロキシアパタイトの結晶が骨芽細胞内のマトリックス小胞内で形成されます。石灰化の第 2 段階では、ヒドロキシアパタイトが膜を通って ECM に分泌され、コラーゲン原線維内に沈着します ( Anderson、1995 )。
骨の機械的伝達
細胞が機械的刺激を生化学的反応に変換するメカニズムは、機械伝達と呼ばれます ( Ingber、2006 )。機械的な力が細胞の挙動に影響を与え、正常な組織の生理機能と疾患の両方において中心的な役割を果たすことは広く受け入れられています。たとえば、内皮細胞は、血液のせん断と圧力の結果として、哺乳動物の組織内で最も大きな力の 1 つ (2 ~ 4 Pa) を受けます。これらの細胞は、流体せん断応力の結果として細胞の形態と方向を変化させ、血管の生理機能と病理を決定することが示されています ( Davies、1995 )。他の例としては、聴覚系や肺などがあります。機械伝達は聴覚の基本であることが示されています。音圧の変化により、内耳の有毛細胞が曲がり、Ca 2+の放出などの生化学シグナルのカスケードが開始されます( Vollrath et al., 2007 ; Gillespie and Müller, 2009 )。肺機能は、組織のひずみ、流体せん断応力、圧縮などの機械的な力によっても制御されます ( Schumacker、2002 )。
同様に、機械的負荷は、骨の質量と構造的適応に対する主な要因の 1 つであることが確認されています。19 世紀に、Julius ( Wolff, 1892 ) は、「骨の変形に関するウォルフの法則」の中で、骨の構造は機械的応力の結果であるという考えを仮定し、それを数学的法則に関連付けました。
流体せん断応力
間質液 (ISF) は体重の主成分 (最大 20%) であり、ECM 全体に分布しています。それは細胞に栄養と老廃物の除去を提供し( Bijlani, 2004 )、組織内の空隙を埋める皮質骨や海綿骨にも存在します。骨の空隙率の 3 つのレベルは次のとおりです:
(1) フォルクマン管およびハバース管 (半径 20 μm) 内の血管空隙率、
(2) 石灰化した骨内のチャネル構造であるラクナ小管系 (LCS)骨細胞とその突起を取り囲む組織(半径 0.1 μm)
(3)鉱物ヒドロキシアパタイトの微結晶とコラーゲン線維の間の小さな空間(半径 0.01 μm)(Cowin and Cardoso、2015)。
ISFの流れは一般にリンパドレナージと関連しており、毛細血管から漏れた血漿は血液循環に戻されます( Swartz and Fleury、2007 )。血液、間質、リンパ管の間の静水圧と浸透圧の違いは、ほとんどの軟組織に存在するゆっくりとした、しかし一定の ISF の原動力であると考えられており、特に安静時の血管の空隙や LCS の小さなチャネルの流れにも影響を与えます。
ほとんどの軟組織における ISF の流れとは対照的に、骨組織では、血圧変化と機械的負荷を引き起こす筋肉の収縮によって、実質的により大きな流量が生成されます (Burger and Klein- Nulend , 1999 ; Knothe Tail et al., 2000年)。機械的負荷により、骨が曲がり、マトリックスが変形します。骨の一方の側では圧縮応力が発生し、もう一方の側では引張応力が発生します 。
結果として生じる ISF 内の圧力勾配により、流体が圧縮領域から張力領域に移動すると考えられています ( Duncan および Turner、1995 )。ISFは、石灰化した骨基質内の小さなスペース、小腔に存在する骨細胞を接続する狭いチャネル、小管を通過しなければなりません。チャネルの寸法が小さいため、血管壁せん断応力に匹敵する高い壁せん断応力が発生します。Weinbaum らによる数値モデル。( 1994 )プロテオグリカンで満たされた小管内の骨細胞プロセスによって感知される機械的負荷によって誘発される FSS は 0.8 ~ 3 Pa の範囲であると推定されました。
高い流体せん断応力への応答
0.5~2 Paの範囲の流体せん断応力(FSS)は、in vitroで骨芽細胞に影響を及ぼし、生化学的要因と遺伝子発現に変化をもたらすことがわかっています。 -FSSは、骨芽細胞培養における細胞内カルシウム、イノシトール三リン酸、一酸化窒素、プロスタグランジンE2、アデノシン三リン酸のレベルを急速に増加させます。 -体液の流れは、オステオポンチン、シクロオキシゲナーゼ-2、c-FOS、コラーゲンIの骨芽細胞遺伝子の発現を調節します。 -ほとんどの実験は、流動時間が短いパラレルプレートフローチャンバー(PPFC)で行われているため、長期間の細胞培養には課題があります。 -骨芽細胞の増殖とマトリックス沈着により、経時的に流路形状が変化し、せん断応力が変化する可能性があります。
低い流体せん断応力への応答
骨芽細胞は、骨形成に重要な役割を果たす骨細胞の一種です。骨基質内に埋め込まれている骨細胞とは異なり、骨芽細胞はより一般的に低い流体せん断応力 (FSS) を受けます。 -体外研究により、骨芽細胞は0.5 Pa未満の低いFSSレベルに反応することが示されています。これらの研究は、平面上で単層状に培養された骨芽細胞を対象に実施されています。 -低FSSに対する骨芽細胞の反応は、高FSSに対する反応と同等であることがわかりました。ただし、低FSSの影響を調査した研究は少ないことに注意することが重要です。 -低FSSは、プロスタグランジンE2 (PGE2)、アルカリホスファターゼ (ALP)、コラーゲンなど、骨芽細胞における特定のシグナル伝達因子の産生を増加させることがわかっています。 -骨芽細胞はまた、流体の流れが少ない条件下ではより増殖する傾向がありました。 -Runx2、ALP、Col I、オステオカルシンなどの骨形成遺伝子のmRNA発現は、低FSS下で増加しました。
まとめ
体外での骨形成には、流体せん断応力が重要な役割を果たします。骨芽細胞などの骨細胞は、体液せん断応力に敏感で、一酸化窒素やプロスタグランジンなどの骨形成シグナル因子を放出することで反応します 。体外モデルを使用して、骨細胞シグナル伝達、コラーゲン沈着、マトリックス石灰化に対する流体の流れの影響を研究してきました。
流体せん断応力が低いと、骨芽細胞の増殖を刺激し、アルカリホスファターゼ活性を高め、フィブロネクチンとフィブロネクチン受容体の発現を高めることができます。ただし、流体せん断応力に対する応答は、加えられる応力の持続時間や大きさなど、特定の実験条件によって異なる場合があります 。全体的に見て、体外での骨形成に対する流体せん断応力の影響を理解することは、改良された組織工学骨インプラントやより効果的な骨疾患モデルの開発に貢献できる可能性があります。
この記事が気に入ったらサポートをしてみませんか?