
遺伝子発現とか(高校理科生物)

★遺伝子発現の仕組み
遺伝子の転写、翻訳が起こり、タンパク質が合成される事を発現という。
細胞にはその機能や構造に応じて必要な遺伝子のみが発現している。
遺伝子の発現もまた様々なタンパク質により調節されている。
・ 真核生物の遺伝子発現
発現は三つの過程から成る。
①転写、②スプライシング、③翻訳である
・遺伝情報の変換(発現の流れ)
① 転写
遺伝子(DNAの塩基配列)が mRNA の塩基配列として写される反応。
遺伝子の情報は傷がつかないように厳重に核内に保管されている。
その為、その情報を使う時も本物ではなくRNAに写したコピーを利用する。
転写されるRNAの量は、一般に発現の高さに比例する。
② スプライシング
イントロンを除去しエキソン同士をつなげる反応。
遺伝子及び mRNA はイントロンとエキソンと呼ばれる領域から成る。
イントロンは翻訳の際に使われない部分で、エキソンが必要な部分である。
核内では作り出すタンパク質に応じて適切なイントロンの除去が行われ成熟した mRNA が生じる。
*選択的スプライシング
スプライシングにおいて使用するエキソンの組み合わせを調節することで、様々なmRNAを作ることができる。
③ 翻訳
mRNA の塩基配列の情報をアミノ酸配列の情報に変換し、タンパク質を合成する反応。
・ セントラルドグマ
遺伝現象に見られる原則のこと。DNAの複製と遺伝子発現の原則。
”半保存的複製”、そして”転写と翻訳による発現”の仕組みが、生物全般に見られる普遍的なルールであるとクリックが予想したため、これをドグマ(教義)と名付けた。またこれは遺伝情報の質に関する方向性とも言える。
複製や発現の際、遺伝情報は DNA から DNA 、 DNA から RNA 、RNA からタンパク質というように、DNA を起点にして利用される。そして、別の物質に情報が写されたり変換されたりするほど、情報の質は下がっていく(壊れやすく、変質しやすくなる)。
そのため、原則として質の良い状態から悪い状態にすることはあっても、逆は行わない。これがセントラルドグマの意味だと考えられる。タンパク質の情報でDNAを合成するようなリスクは負わないと言うことである。
ただし、RNAからDNAを合成する例外は存在する。これはレトロウィルスなどの一部のウイルスが行う逆転写である。これによりそれらのウイルスは変異しやすいというリスクを負うが、宿主の体内で免疫から逃れるためにはそれが有利に働くことが場合があるのだろう。
もしくは進化的にRNAを遺伝情報として利用することが普通で、そこから変化してセントラルドグマのようなDNAを遺伝情報として使用するグループが生じたのかもしれない。このような後者の考え方はRNAワールド仮説と呼ばれている。
・ レトロウイルス(HIVなど)
逆転写酵素を利用するRNAウイルス(遺伝子の本体がRNAであるウイルスを、RNAウイルスと呼ぶ)。
感染すると逆転写酵素を用いてRNAからDNAを合成し、それを宿主のDNAの一部にする。そして自身の遺伝子を宿主に合成させて増殖していく。
DNAより質の悪いRNAを遺伝子の本体とするため突然変異が起きやすく、遺伝情報が不安定である。逆転写の発見者であるテミン、ボルティモアらはノーベル賞を受賞した。
●RNAの種類
転写、翻訳の際には3種類のRNAが機能する。
活躍する順番に ⑴ mRNA、⑵ tRNA、⑶ rRNAである。
⑴は遺伝情報のコピーである。
⑵は核酸の情報をタンパク質の情報に変換するアダプターとして機能する。
⑶はリボソームのパーツであり、これと mRNA の相互作用を支えている。
●遺伝暗号(コドン)
アミノ酸や読み枠の開始点、終了点を決める塩基配列の3つ組。
DNAやRNAでは遺伝情報が塩基配列(4つの文字から成る文字配列)として存在する。これがタンパク質の設計図としてタンパク質のアミノ酸配列を決定する。
では、例えば「TACCCGTTTGGG」 というDNAの塩基配列はどのようにタンパク質の構成(アミノ酸の配列)を決めているのだろうか。
ここで転写を考える。そこでは相補性が利用されている。これは塩基配列の情報を正確に伝える、変換する時に適した性質である。従って、翻訳でも相補性を利用してアミノ酸配列を決定していく。つまり、mRNAの塩基配列を相補性によりアミノ酸配列に変換する。
この変換の仕組みの理解は一種の暗号解読である。なぜなら塩基配列をアミノ酸に対応させることは、英語を日本語に変化するのと同じだからである。どんな文法(ルール)と単語の対応があるのかを理解することが重要である。
ルールと単語は次のようなものである。
「アミノ酸や読み枠の情報(どこから読み取りが開始されるかなど)」は、塩基配列における「連続した3つ組の組み合わせ」によって決まる。
つまり、3つ組の塩基配列が重要で、塩基3つごとに1つのアミノ酸が対応するというルールで、かつその3つの塩基が配列ごとに違ったアミノ酸を意味するのである。すなわち単語として機能するのである。
よって、DNAではこのような3つ組の塩基配列をトリプレット、mRNAでは遺伝暗号(コドン)と読んで区別する。また、翻訳で見られるコドンの規則性を表にしたもの、言わば塩基配列をアミノ酸配列に変換するための辞書を、遺伝暗号表(コドン表)という。
従って、塩基配列における相補性と遺伝暗号表を使えば、DNAの塩基配列さえわかるとその解読が可能であり、TACCCGTTTGGG は、mRNAで AUG GGC AAA CCC になり、翻訳後にメチオニン-グリシン-リシン-プロリンとなる事が分かる(コドン表で確かめよう)。
●遺伝暗号の解読
ニーレンバーグ、コラーナの実験により、コドンは解明された。
・ ニーレンバーグの実験
大腸菌の内部で起きる反応が知りたい時、小さな大腸菌を調べるよりも、内部のシステムを予め取り出し研究する方が都合がよい。そこで、ニーレンバーグは大腸菌の抽出液を使って、試験管内でタンパク質合成が行えるようにした。RNAを加えるだけでタンパク質ができるシステムだ。
そして、生体での発現が再現されているかを確認するため、自然界には無いポリウリジル酸(ウラシルのみから成るRNA)を使って、検証(対照実験)を行った。発現の仕組みが再現されていれば、自然界に無いRNAからのタンパク質の合成は起こらないだろうと考えたのである。
しかし、実際にはフェニルアラニンから成るタンパク質が合成された。彼の実験の条件では、細胞内部とは異なりランダムな位置から無理にmRNAの翻訳が行われていたのである。だが、その一方でこの失敗は人工的なRNAを利用したタンパク質合成を可能にしたのである。
そして、これを利用することによりさらにコドンの解明も一挙に進むことになる。なぜなら、この実験からUUUというコドンがフェニルアラニンであることが判明したのである。また、用意する人工的なRNAを調べたい任意の配列にすることで、それがどんなアミノ酸に対応するかを次々に調べることが可能になったのである。
・コラーナの実験
コラーナは簡単な繰り返し構造を持つ人工RNAの合成に成功し、ニーレンバーグの実験に応用した。AUAU…、GCGC…等の塩基配列を持つRNAを用いてどのようなタンパク質ができるかを調べたのである。その結果、またたく間にコドン表が完成したのである。
・ 開始コドン
AUG
・ 終始コドン
UAA、UGA、UAG
●分泌タンパク質やリソソームタンパク質の輸送
① 粗面小胞体で合成された分泌タンパク質などは、
小胞体内部で組み立てや品質検査を受ける。
② 目的地に運ぶため、小胞が形成される(袋詰め)。
この時、生体膜表面では被覆タンパク質が働く。
③ 細胞骨格を利用して、分子モーターが小胞を輸送。
④ 目的地の膜と小胞の膜が融合し、荷物が届く。
・ エクソサイトーシスとエンドサイトーシス
エクソは「外へ」、エンドは「内へ」という意味である。
サイトーシスは「細胞の活動」ということである。
つまり、細胞外への分泌と細胞内への取り込みを表す、
●大腸菌の転写と翻訳
大腸菌(原核生物)には核膜という構造やスプライシングという機構が存在せず、転写と翻訳が時間的、空間的に同時に進行する。これは電子顕微鏡による観察から明らかにされた。初期の分子生物学の偉大な成果の1つである。
そのため、その電顕の模式図を元にした定番の問題がある。歴史的に重要な研究が背後にあると科学者としての資質が問えるとの考えがあるのかもしれないと思う。あと気分的に楽しくなるかも。
問題を解く際は、電顕の模式図や画像から、
①RNAポリメラーゼの移動する向き、
②DNAとRNAの方向性の違い、
③リボソームの移動する向き
などが問われ、mRNAやタンパク質の長さから、それらを判断する。
(現象を動的にイメージする能力は大事であると思う)
考えるポイントは、
① 転写を先に始めたRNAポリメラーゼほど、長いmRNAをぶら下げる。
② 多くのRNAポリメラーゼが連続して転写を行っている部分を見る。
①より長いmRNAの方向にポリメラーゼが進んでいるとわかる。
③ mRNAの5’末端は最初に転写される。この部分から翻訳が開始される。
④ リボソームも5’から3‘方向に翻訳を行う。
よって反応が進むほど長いポリペプチドがリボソームにぶら下がる。
●遺伝子とタンパク質
遺伝子は多くの場合タンパク質の設計図として機能する。
では、これはどのように明らかになったのだろうか?
キーワードは「異常」と「変異」である。
遺伝の研究には異常な生物がよく使われる。
これは、異常な形質と遺伝子の破壊が強く関連するからである。
生物は沢山の遺伝子を組み合わせて体というシステムを作る。その為、突然変異によって何らかの遺伝子が壊れると、それが引き金となってシステムが破綻し、大きな変化が生じる場合があるのだ。
言うなれば、作るのは大変で壊すのは簡単なのである。
とはいえ、形質が大きく変化した変異体では発生において重要な遺伝子が壊れている場合が多い。
このような異常な生物を使うと、遺伝子が壊れた結果としてどのようにして形質に違いが出るかを分子レベルで上手く比較することができる。特に20世紀初頭は医学や生化学が発達し、人工的に突然変異を誘発する技術すら生まれた。そのため、それらの変異体を正常型(野生型)と分子レベルで比較することが可能になったのである。
実際に、ヒトも含めて様々な生物で遺伝性の病気や突然変異体が調べられていった結果、遺伝子の破綻と形質が遂には分子レベルで結びついたのだ。
遺伝子が壊れると、多くの場合にタンパク質が異常になることが示されたのだ。
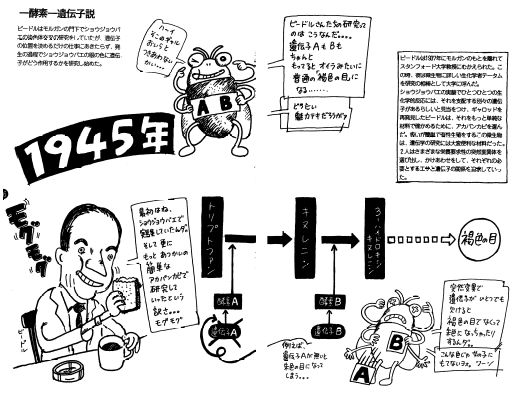
・ 一遺伝子一酵素説
ビードルとテータムはアカパンカビの突然変異体から栄養要求株(突然変異体の一種)を分離し、特にアルギニン要求株(アルギニンが与えられなければ死ぬ)を詳しく調べた。
その結果、これらがアルギニンの合成に関わる酵素のいずれかを失っていると結論した。つまり、これらの変異体では遺伝子が壊れ、その結果として酵素が壊れていると結論した。
・ ビードルとテータムの実験
① アルギニン要求株(メンデルの法則に従う)を詳しく調べ、
次の三つに分類できることを発見した。
⑴アルギニンを与えた場合にのみ生育可能。
⑵アルギニンに似たアミノ酸(シトルリン)を与えた場合でも生育可能。
⑶シトルリンに加え、
オルニチン(シトルリンに似たアミノ酸)を与えた場合でも生育可能。
② アルギニンとシトルリン、シトルリンとオルニチンは良く似ている。
従って、アルギニン様の2つのアミノ酸は、これを作る為の材料、
つまり代謝過程の中間体だと考えられた。
③ ②が正しければ、次のような推論が可能となる。
⑴はどのような材料が与えられても、アルギニンが作れない株
⑵はシトルリンからアルギニンを合成できる株
⑶はシトルリン、オルニチンからアルギニンを合成できる株
④ ⑴のように材料を与えても代謝できないというのは、実験、料理でいうと
器具や装置が壊れていて作れないということであろう。
つまり、代謝の場合で考えると酵素という装置が壊れているはずである。
⑤ ④のように考えると、①はアルギニンの代謝に必要な複数の酵素の内、
どれか一つが欠けたものと解釈できる。
つまり、各変異体では代謝に関わる酵素がそれぞれ壊れていると考える。
これが一遺伝子一酵素説と呼ばれる考え方でる。
(各遺伝子は異なる酵素の設計図であるという仮説である)
⑥ 酵素は基質特異性を持つため、各反応ごとに1つの酵素が必要になる。
ここで、一遺伝子一酵素説を使って3つの変異体を考えてみる。
すると、⑴~⑶は次のように解釈できる。
⑴ アルギニンを作るための最後の仕上げの酵素がない。
そのため、生育にはアルギニンが必須となる。
⑵ シトルリンからアルギニンが作れる。
つまり、シトルリンをアルギニンに変える酵素を持つ。
しかし、シトルリンを作るための酵素が無い。
だから、シトルリンかアルギニンが生育に必要となる。
⑶ オルニチンやシトルリンからアルギニンを作れる。
オルニチンからシトルリンを作る酵素があるということだ。
だが、オルニチンを作る酵素が無いと考えられる。
それゆえ、3つの内のアミノ酸のいずれかが生育に必要になる。
⑦ 一遺伝子一酵素説はその後、一遺伝子一ポリペプチド説に発展した。
これは4次構造をとるタンパク質やコラーゲン等、様々な変異体の研究で
酵素ではないタンパク質の異常が発見されたためである。
・ 酵素の遺伝子と遺伝病
一遺伝子一酵素説が当てはまる例として、フェニルケトン尿症やアルカプトン尿症がある。
前者はフェニルアラニンをチロシンに変える酵素が異常で、フェニルケトンが増加し乳児期には脳の発達を阻害する。
「アルジャーノンに花束を」の原作では主人公がこれであった。
後者は、アルカプトン(フェニルアラニンやチロシンの代謝で作られる物質)が壊れた酵素で分解できずに尿中に増える病気である。
尿が黒くなるほか、心臓や関節、腎臓などに障害が出る。
この記事が気に入ったらサポートをしてみませんか?