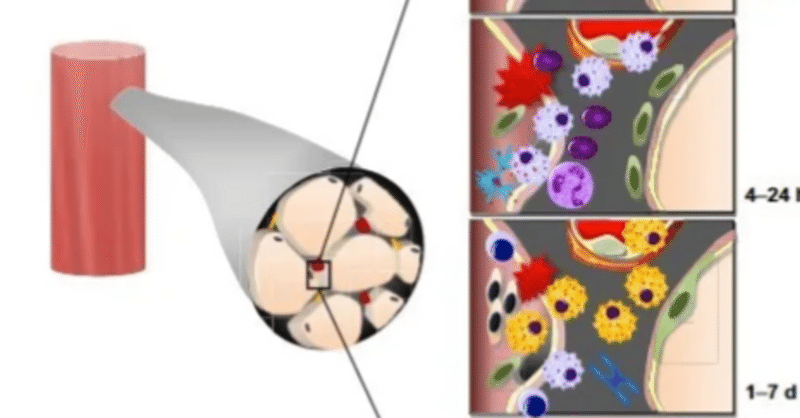
運動誘発性筋損傷後に生じる適応プロセス

運動誘発性筋損傷(Exercise-Induced Muscle Damage、EIMD)は、30年以上にわたり、運動とスポーツ科学の研究で重要な焦点となっています。これは一時的な超微細構造の筋原線維の破壊、筋力とパワーの喪失、遅発性筋痛(DOMS)、腫れ、影響を受けた肢の可動域の減少、筋細胞内の酵素やタンパク質(例:クレアチンキナーゼ(CK)、ミオグロビン)の全身性流出などに特徴づけられる状態です。運動誘発性筋損傷に関連する機械的な変化と代謝ストレスは、骨格筋を構成するさまざまな細胞タイプを刺激し、その後の組織修復とリモデリングを開始します。具体的には、衛星細胞(筋幹細胞)、炎症細胞(例:好中球、マクロファージ、Tリンパ球、マスト細胞)、血管細胞(例:ペリサイト、内皮細胞)および基質細胞(例:線維芽細胞)が骨格筋の細胞外マトリックス(ECM)内で互いに相互作用します。これらの細胞間相互作用のダイナミクスは、筋損傷からの回復の効果と経過時間を決定します。
筋損傷と炎症のプロセスについては他の場所で広くレビューされています。
運動誘発性筋損傷の基本的なメカニズム、特徴、および経時的な変化。
運動誘発性筋損傷からの回復中の筋内炎症の経時的な変化と役割。
運動誘発性筋損傷および筋内炎症に対する効果的な治療法。
運動誘発性筋損傷の基礎
筋肉の超微細構造の破壊、筋力の低下、遅発性筋痛(DOMS)、および筋酵素の流出が、収縮が遠心的(すなわち伸長)であれば求心性収縮(短縮)の場合に比べて大きく、これらの指標の回復が遅いことはよく受け入れられています。収縮が求心的な場合は運動誘発性筋損傷は発生しないが、長い筋の長さでの等尺性収縮および遠心的収縮では、低強度であっても運動誘発性筋損傷が明らかになる。遠心的な運動の後の筋力低下は、運動誘発性筋損傷の最良の指標と考えられており、そのメカニズムは次の理論モデルで概説されています。遠心運動中の機械的なひずみは、半分のサルコメアの不均一性とフィラメントの重なりを超えたサルコメアの過度な伸張を引き起こし、「弾けたサルコメア」をもたらします。これらの変更は直接的に力の生産を減少させ、サルコレンマおよびt管構造に過負荷をかけます。これにより、ストレッチ活性化チャネルの開口、膜の破壊、および興奮-収縮結合の機能不全が引き起こされます。ストレッチ活性化チャネルおよび/またはサルコレンマの透過性部分を介して細胞質に入るCa2は、カルパイン酵素の刺激により収縮タンパク質または興奮-収縮結合タンパク質を分解し、筋力の長期間の低下を引き起こす可能性があります。 DOMSも筋損傷の一般的な症状ですが、DOMSの正確なメカニズムは依然として不確かな部分があります。一般的には、筋線維の微小損傷とそれに続く炎症がDOMSを引き起こすと考えられています。ただし、ラットの筋肉では、明らかな微視的な筋肉の損傷や炎症の兆候がないにもかかわらず、遠心的な筋収縮の1〜3日後に機械的な痛覚過敏が発生します。遠心的な筋収縮の後、機械的な痛覚過敏を引き起こす2つの経路が関与しています。
1)B2-ブラジキニン受容体-神経成長因子経路の活性化、
2)COX-2-神経栄養因子経路の活性化。
これらの神経栄養因子は筋線維および/または衛星細胞によって産生される可能性があります。これらの物質は筋感覚受容体を刺激することによって直接DOMSを引き起こす可能性があります。あるいは、細胞外受容体に結合し、筋線維から神経栄養因子の分泌を誘導して感覚神経受容器の刺激とDOMSを引き起こす可能性があります。 DOMSは、筋線維の損傷と炎症よりも、細胞外マトリックス(ECM)の炎症と関連していると考えられています。
運動による筋のサルコメア、膜、およびECM構造の破壊の生物学的な重要性は未だに議論の的となっています。Zディスクの流れ、筋束間のペリミセアル領域の広がり、および筋束内での筋線維の分離といった初期の形態学的観察は、筋損傷の証拠と解釈されました。しかし、より現代的な理論では、遠心運動自体が筋線維を損傷させるのではなく、Zディスクの流れ、および破壊はむしろ筋のリモデリングと適応を表している可能性があると提唱されています。これらの超微細構造の変化が具体的な生物学的意義を持っているかどうかにかかわらず、これらは運動後の筋肉の変化に敏感であり、機械的な負荷/収縮モードの違いと筋機能の変化に関連しています。
運動誘発性筋損傷の時間経過
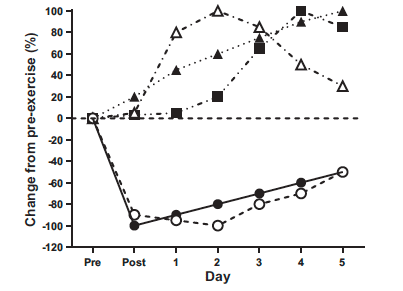
遠心運動直後には通常の筋原線維のバンディングパターンの破壊が見られる筋線維の数が増加します。Zディスクとサルコメアの破壊は、運動後1〜3日の間にピークを迎えるようですが、運動後6〜8日まで持続することがあります。運動後の筋力の喪失の程度と、筋力を正常に戻すためにかかる時間との間には時間的な関連があります。運動直後に筋力が20%減少すると、通常は2日以内に回復します。対照的に、運動直後に筋力が50%以上減少する場合、特に初回の遠心的な筋収縮に対する初期の暴露の場合、運動後7日まで筋力が運動前の値を下回ったままです。激しい遠心運動の後の数日間における筋力、可動域、DOMS、肢の周囲囲(腫れ)、および血中CK活性の変化の時間経過は異なります。筋力の回復が長引く場合でも、DOMSは運動後約4日で解消されます。筋腫脹は運動後4〜5日でピークに達し、筋損傷の血中マーカーの増加も遅れます。筋力の変化は、運動誘発性筋損傷の他のマーカーの変化の大きさと時間経過に影響を与えるようです。
運動誘発性筋損傷からの回復に影響を与える要因
運動誘発性筋損傷からの回復に影響を与える最もよく知られている要因は、前回の筋損傷です。初回の筋損傷性のある運動の後、筋肉は適応し、症状が軽減され、次回の運動後にはより迅速に正常に戻ります。この現象は「繰り返し運動効果」として知られています。これらの保護効果は、低強度の遠心筋収縮、または長い筋の長さでの最大等尺性収縮によって生じ、これらは運動誘発性筋損傷の症状を引き起こさないか(またはわずか)もしくはより軽微に引き起こします。繰り返し運動効果は、対側の筋肉にも与えられ、対側の腕による初回の運動に比べて対側の筋誘発性筋損傷が少なくなることがあります。
高い筋トルクで行われる運動、増加する筋収縮回数、および筋長の長い筋での運動では、運動誘発性筋損傷が増大し、または回復が遅くなる可能性があります。単一関節と複数関節を使用した運動では、前者の方が後者よりも運動誘発性筋損傷が大きい可能性があります。
腕と脚、および膝屈筋と膝伸筋を使用した運動では、いくつかの条件下で運動誘発性筋損傷が増大する可能性があります。
収縮速度は30回の筋収縮後に筋力に影響を与えませんが、210回の筋収縮後、高速(210°/s)対低速(30°/s)での収縮では筋力の喪失が大きくなります。男女間での運動誘発性筋損傷からの回復が異なるかは不明であり、これは研究参加者の年齢やトレーニング状態、運動プロトコルの種類と強度のバリエーションが影響している可能性があります。ただし、性差が大きいわけではないようです。
運動誘発性筋損傷からの回復には、反復とセットの構成、筋収縮のセット間の休息間隔、または損傷した筋肉での筋収縮の実行は影響を与えないようです。
高い筋収縮と低い筋収縮とを比較した場合、おそらく高い筋収縮や増加した収縮回数に関連するより大きな機械的な負荷は、契約タンパク質とECM(結合組織)により大きな損傷を引き起こし、それによりより重度の運動誘発性筋損傷が生じる可能性があります。
筋長の長い筋で行われた筋収縮は、おそらくサルコメアの長さの不均一性の度合いが大きく、伸張された、より弱いサルコメアとECMの大きな混乱を引き起こし、それによりより重度の運動誘発性筋損傷が生じる可能性があります。単関節の運動と比較して多関節の運動後に運動誘発性筋損傷がより大きいのは、おそらくより小さくて弱い筋肉グループの少ない活動と、これらが過伸展に対してより脆弱であるためです。神経制御は筋収縮と求心性収縮または遠心性収縮の間で異なり、非訓練者は通常、最大筋収縮中に筋を完全に活性化できないことがあります。また、筋収縮中の運動単位放電率は、主に脊髄の興奮性が低下するため、等尺性収縮と比較して低いです。
腕対脚、および膝屈曲対膝伸展の運動後のより大きな運動誘発性筋損傷は、おそらくこれらの筋グループにおける規則的な機械的負荷の差に起因する可能性があります。
多数の早い速度の筋収縮に続く運動誘発性筋損傷の差は、2つの理由から生じる可能性があります。まず、早い収縮は長い長さでより大きな力を生み出す可能性があり、これによりタンパク質への損傷リスクが増加します。第二に、早い収縮は力を生み出すことができる交差橋の数が少なくなる可能性があり、これによりアクティブな交差橋ごとの機械的ストレスが増加するかもしれません。この効果は、速筋が収縮回数が増加するにつれて悪化する可能性があります。
急性応答と運動誘発性筋損傷後の炎症の解消
用語「炎症」は、しばしば具体的な定義やそれが「良い」プロセスなのか「悪い」プロセスなのかといった価値判断なしに広く使用されます。スポーツ医学の文脈では、「炎症」は怪我を受けた組織内での臨床的、生理的、細胞、および分子レベルの変化を包括しています。歴史的に、運動誘発性筋損傷後の筋肉の炎症は、時折、組織損傷、痛み、および遅れた回復と関連する有害なプロセスと見なされることがありました。しかし、この広い視点は炎症の多様な側面を考慮に入れていません。炎症が筋肉の修復と再生の基本的なプロセスであるという考えが、今では受け入れられつつあります。非病態学的条件(例:運動誘発性筋損傷後)では、筋内炎症は緊密に調整されたダイナミックなプロセスであり、最終的に適応的なリモデリングとホメオスタシスへの回帰につながります。
組織の組織学的証拠と筋肉中のサイトカインmRNAおよびタンパク質の量の定量に基づいて、ヒトにおける運動誘発性筋損傷後の筋肉の炎症にあります。また、これらの調査は、筋肉内炎症の調節とその結果に関する重要な知見を提供するため、主要な動物および細胞培養研究も参照しています。筋肉と免疫の相互作用の動態は筋肉の機能的な回復にとって重要な要素であるため、収縮運動後の筋肉の炎症の経時的な経過にも焦点を当てています。
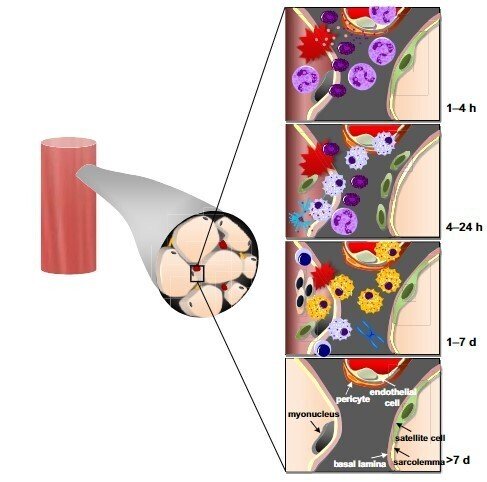
運動後のヒトの筋肉炎症の経時的な経過
炎症細胞(白血球)の筋肉組織への蓄積は、組織学的な観察によって同定され、運動誘発性筋損傷の基本的な兆候と考えられています。さまざまな種類の「筋肉損傷」運動を含む人間の研究では、白血球が筋肉組織に蓄積する証拠が提供されています。白血球は、激しい、大容量の、または慣れていない抵抗運動、下り坂のランニング、およびランニングを含む長距離走行/超耐久運動の後の筋生検サンプルで観察されています。局所の炎症反応が運動ではなく反復された筋生検によるものかどうかについては議論がありますが、生検手順自体から生じる炎症が運動から生じる炎症と比較して小さなものであるという証拠もあります。
白血球は運動後すぐに筋肉に蓄積し始める可能性があります。放射線標識された白血球、主に筋肉組織の微小血管に存在する好中球が、運動後1から24時間の間に観察されています。組織学的な検査では、通常、白血球が運動後24〜48時間の間に筋肉の細胞間空間に蓄積することが示されます。好中球の蓄積に関する具体的な証拠は24時間を超えて制限されています。これは、好中球を検出する方法論的な難しさや、より可能性としては、好中球が再生筋肉から迅速に消失するためかもしれません。ヒトの骨格筋では、回復の後の時点である48時間から7日以上にわたって、モノサイト/マクロファージの数がより一貫して増加しています。同じ回復時点で、一部の個体は筋肉中に大量の白血球を示し、他の個体は非常に少ない白血球を示すことがあります。
これらの観察結果は、運動誘発性筋損傷に続く好中球とモノサイトが循環に動員されるという考えを支持しています。その後、これらの細胞は筋肉に移動し、ファゴサイトーシスとプロテアーゼ酵素(例:エラスターゼ、ミエロペルオキシダーゼ)、反応性酸素および窒素種を放出して損傷した筋肉組織を分解します。以前の研究では、中程度の筋肉損傷を伴う持久性運動(サイクリングに続くランニング)の後、血清ミオグロビン濃度とCK活性の上昇を示して、骨格筋のトランスクリプトームの変化の時間経過を調査しました。これらのデータは、運動後3時間での早期の白血球の筋肉への移動と免疫活性化を示唆しています。さらに、白血球の存在、免疫関連のシグナリング、筋肉内ECMの適応的なリモデリングに関連する機能的な転写活性は、運動後96時間まで明らかでした。
組織内在のマクロファージなどの白血球も、運動後に血液由来のモノサイトの募集および蓄積とは別に(またはそれがなくても)、活性化される可能性があります。特に、激しい運動誘発性筋損傷(後述)の約4-7日後、白血球は運動した筋肉組織の細胞内空間にも侵入します。強烈な意図的な筋収縮に対しても、重度の筋線維壊死が発生する証拠はほとんどありません。筋肉が電気的に刺激された収縮に続いても、重要な壊死が発生します。筋肉収縮中の筋線維のリクルートメントのパターンは、したがって壊死の程度を決定するかもしれません。あるいは、全体の筋線維に影響を与えずにセグメンタルな筋線維の壊死が発生する可能性もあります。このような「重度」なケースでは、白血球は運動後3週間経過しても筋肉組織中に観察されています。
組織学的な白血球浸潤の証拠に加えて、運動誘発性筋損傷は筋肉中のサイトカイン/ケモカインmRNAおよびタンパク質の発現の増加とも関連しています。特に、IL-6、C-X-Cモチーフリガンド8(CXCL8;またの名をIL-8)、C-Cモチーフケモカインリガンド2(CCL2;またの名をモノサイトケモタクチックプロテイン-1としても知られる)のmRNA発現の変化に焦点が当てられており、主に2つの時間枠、すなわち運動後1〜4時間および24時間後に行われています。mRNA発現と比較して、運動後の筋肉でのサイトカイン/ケモカインタンパク質発現の変化についてはほとんど知られていません。
筋肉負荷が筋肉の炎症に及ぼす影響と筋肉の機能回復との関連
ヒトの研究では、筋肉に白血球が蓄積することで示されるように、筋肉の炎症は筋肉の損傷の程度に応じて徐々に進行することが示されています。 -力を生み出す能力または筋力によって測定される筋肉の機能は、筋肉損傷の信頼できる指標と考えられています。 -筋力の変化を測定し、筋生検サンプル中の白血球の存在を分析した研究により、筋機能と白血球蓄積との間に関連があることが判明しています。 -筋力の低下と完全な筋力回復に必要な時間は、筋肉の損傷と回復の指標として使用されます。 -「軽度」の運動による筋肉損傷(筋肉機能が 20% 未満低下し、2日以内に完全に回復する)に反応して白血球が蓄積される証拠は限られています。 -しかし、研究によると、2日以内に筋肉機能が回復したとしても、最大偏心筋収縮後も筋肉の筋内膜および筋膜周囲領域にCD68+マクロファージが蓄積する可能性があります。 -一方、一部の研究では、遅発性筋肉痛(DOMS)があり、血液顆粒球と血清クレアチンキナーゼ(CK)活性が増加しているにもかかわらず、下り坂を走ってから48時間後に白血球炎症の兆候がないことが報告されています。 -これは、筋肉損傷の一般的な症状であるDOMSが、必ずしも筋線維間の白血球の蓄積に関連しているとは限らないことを示唆しています。
中程度の運動誘発性の筋肉損傷(筋力が 20% 以上低下し、7日以内に回復する)は、筋肉への白血球の蓄積に関連していることがわかりました。 -筋線維への白血球の細胞内浸潤は、通常、「ハイレスポンダー」と呼ばれる、筋肉機能の最も顕著で長期にわたる低下を示した個人で観察されました。 -力の生成能力が 50% 以上低下し、回復期間が1週間未満であることを特徴とする重度の運動誘発性筋肉損傷に関する研究でも、筋肉に白血球が蓄積したことが報告されています。 -筋肉中の白血球数が最も多かったのは、分節性筋線維壊死(筋線維の死)の徴候があったのと同じ時点で観察された。 -全体として、広い可動域にわたる最大限の偏心運動によって誘発される中程度から重度の筋肉損傷に反応して、筋肉への白血球の蓄積が一貫して観察される。
筋肉の再生/回復中の筋肉と免疫の相互作用の根底にある白血球の機能とメカニズム
白血球は、免疫反応に重要な役割を果たす機能の一種です。 -筋肉の再生と回復の過程では、複数の種類の免疫細胞が筋肉組織と相互作用し、筋肉の修復と再生のさまざまな段階に不可欠です。 -げっ歯類を対象とした研究により、筋肉の修復と再生に免疫細胞が関与していることを示す証拠が増えています。 -筋肉損傷に対する初期反応は、主に好中球と炎症誘発性マクロファージの蓄積によって引き起こされる炎症誘発性反応を特徴としています。 -白血球の一種である好中球は筋肉損傷の一因となり、マウスの収縮誘発性損傷による筋肉損傷後の筋肉リモデリングと機能回復を損なう可能性があります。 -しかし、好中球をマクロファージと共培養すると、マクロファージの細胞毒性と筋細胞溶解能力が低下します。 -これは、好中球がマクロファージの細胞毒性を変化させ、その結果、筋細胞の溶解に必要なマクロファージが少なくなる可能性があることを示唆しています。 -これらの知見は、筋肉損傷の炎症誘発性段階で好中球とマクロファージの間に相互作用があることを示している。
炎症誘発性マクロファージの特徴
炎症誘発性マクロファージは、CD68と呼ばれる細胞表面分子を強く発現する特定の種類のマクロファージです。 -これらのマクロファージは伝統的にM1マクロファージと呼ばれてきました。 -ただし、これらのマクロファージは、受容体の発現パターンに基づいてさらに特徴付けられています。具体的には、Gr1 high または Ly6C pos CX3CR1 lo マクロファージと呼ばれています。 -これらの受容体の発現は、炎症誘発性マクロファージのさまざまなサブタイプを同定して区別するのに役立ちます。 -好中球とM1/Ly6C pos CX3CR1 lo マクロファージはどちらも、食作用や活性種生産などのプロセスを通じて細胞破片を除去する上で重要な役割を果たします。 -ただし、これらの炎症細胞による反応性種の過剰産生は、筋肉の損傷を悪化させる可能性があります。
筋肉の再生とリモデリングの妨げ -特定の筋肉炎症反応を妨害すると、筋肉の再生とその後の適応的リモデリングに影響を与える可能性があります。 -例えば、筋肉損傷の前にマウスの好中球を枯渇させると、おそらく好中球によるマクロファージの動員障害が原因で、筋再生が損なわれる。 -炎症誘発期の初期段階で、傷ついた筋肉への単球 (免疫細胞の一種) の動員を阻止したり、傷ついた筋肉を抗炎症性サイトカイン IL-10 で治療したりすると、筋肉の再生も損なわれます。
筋肉再生中のマクロファージの表現型可塑性 -マクロファージは、筋肉の再生中に顕著で動的な表現型の可塑性を示します。 -つまり、組織の環境によって特徴や機能が変化する可能性があるということです。 -特に、炎症が解消する過程で、表現型が炎症誘発性M1マクロファージから抗炎症性M2マクロファージにシフトします。 -炎症誘発性反応から抗炎症反応へのこの移行は、炎症の解消と筋肉再生の促進にとって重要です。
筋肉の損傷と炎症からの回復を促進するための戦略
運動による筋肉の損傷は、筋肉が慣れていない遠心性(伸長)収縮にさらされると発生します。その結果、筋肉組織の構造が変化し、筋力の低下、痛みや腫れの増加、筋肉細胞からのタンパク質の放出などのさまざまな臨床症状や症状が起こります。 -筋肉がこの損傷から回復するのにかかる時間は、最初の損傷の程度によって異なります。初期損傷の程度は、運動の強度と持続時間、関節の角度/筋肉の長さ、運動中に使用される特定の筋肉群などの要因の影響を受けます。 -これらの要因が筋力、痛み、腫れに及ぼす影響はよくわかっていますが、筋肉内炎症や筋肉の適応とリモデリングの分子的側面にどのように影響するかについての知識はまだ限られています。 -炎症は従来、回復過程に有害であると見なされてきましたが、現在では、炎症反応の制御が筋肉の修復と再生に重要であることが認識されています。 -動物実験では、好中球、マクロファージ、肥満細胞、リンパ球、線維脂肪形成前駆細胞、周皮細胞など、さまざまな種類の細胞が筋肉組織の再生を促進する役割を果たすことが示されています。ただし、これらの細胞が運動による筋肉損傷に反応するかどうかを判断するには、さらに研究が必要です。 -運動による筋肉損傷の兆候や症状を軽減する上で、理学療法、薬物療法、栄養などのさまざまな介入の有効性を調査する研究が数多く行われています。しかし、これらの研究の結果はまちまちであり、これらの治療法が回復過程における炎症と筋肉のリモデリングにどのように影響するかを理解するには、さらなる研究が必要です。
筋回復の治療
多くの治療法がテストされ、運動後の筋機能の回復と筋肉痛の軽減に効果があることが確認されています。 -マッサージ、冷水浸漬、着圧着などの一部のトリートメントでは、筋肉の回復を改善する効果が一貫して示されています。 -ただし、他のほとんどの治療法では、身体的効果という点で一貫して肯定的な結果が得られていません。 -治療によっては、身体的な効果を裏付けるエビデンスが不足している場合でも、運動の回復に重要な知覚効果がある場合があることに注意することが重要です。 -例えば、これらの治療は、構造リモデリングが起こる前に、痛みの知覚を隠したり、回復を早めたりすることがあります。 -しかし、これらの治療の中には、実際にはさらなる筋肉損傷のリスクを高めるものがあるという懸念があります。
運動後の回復戦略が筋肉内炎症に及ぼす影響を考えることは重要です。
また、運動後の回復治療に対する反応の男女間の類似点または相違点を理解することです。 -超微細構造損傷(Zラインストリーミングなど)と筋原線維損傷(壊死など)との関係も重要です。 -筋肉の細胞外マトリックス (ECM) 内で炎症を引き起こす機械的および生化学的要因を理解することが重要です。 ECM内の炎症が、遅発性筋肉痛 (DOMS)、筋力低下、腫れなどの筋肉損傷の症状に関連しているかどうか、またどのように関連しているかを調査する必要があります。ダメージを与えない運動の後に筋肉と免疫の相互作用が起こるかどうかを判断することが重要です。
この記事が気に入ったらサポートをしてみませんか?